1. Definition
Unter dem Begriff des Lobus limbicus wurden von P. Broca verschiedene Strukturen der mittleren und basalen Großhirnoberfläche zusammengefaßt, die sich wie ein Saum (Limbus) um den Hirnstamm legen. Unter Einbeziehung der subkortikalen Areale, die mit dem Lobus limbicus in Verbindung stehen, wurde dieser Begriff später zum "Limbischen System" erweitert.
Tabelle 14-1: Bestandteile des Limbischen Systems.
| Kortikale Bestandteile (Pallium) |
Allocortex =Archicortex Peri-Allocortex=Peri-Archicortex |
Hippocampusformation (Ammonshorn, Gyrus dentatus, Subiculum) Gyrus cinguli, Gyrus parahippocampalis |
| Subkortikale Bestandteile (Subpallium) |
Mandelkern Limbische Vorderhirnregion (Area septi, Ncl. interstitialis der Stria terminalis, Ncl. accumbens) Limbische Areale im Zwischenhirn (Thalamus, Hypothalamus, Ncl. habenularis) Limbische Mittelhirnregion (Ncl. interpeduncularis, Area tegmenti ventralis, Ncl. tegmenti dorsalis) |
|
Abb. 14-1: Kortikale und subkortikale Strukturen des Limbischen Systems

Erklärungen:
Kortexabschnitte:
1: Area parolfactoria,
2: Gyrus subcallosus,
3: G. cinguli,
4: Isthmus cinguli,
5: G. parahippocampalis,
6: Hippocampus-formation (die rudimentären Strukturen sind nicht dargestellt). Seine Assoziationsfasern bilden das Cingulum und den Fornix. Subkortikale Strukturen:
7: Mandelkernkomplex ,
8: Area septi,
9: Cp. mamillare,
10: ventrale amygdalofugale Verbindung,
11: Lamina terminalis
Legende
Der limbische Kortex (Lobus limbicus) gehört zum Archicortex und liegt an der medialen Oberfläche der Hemisphären ("Innenhirn"). Er bildet dort einen Ring um den Balken und den Hirnstamm.
2. Kortikale Bestandteile
Der Allocortex (Archicortex) umfaßt den Hippocampus selbst (Ammonshorn, Seepferdchen), den Gyrus dentatus und das Subiculum. Er grenzt medial an den Balken und den Hirnstamm; seitlich, im Übergangsbereich zum Isocortex liegt der Periallocortex (= Periarchicortex, =Mesocortex). Dieser umfaßt Gyrus cinguli und Gyrus parahippocampalis. Den vorderen Abschnitt des Gyrus parahippocampalis bildet der Cortex entorhinalis.
2.1 Peri-Archicortex
= Mesocortex: Gyrus cinguli, Gyrus parahippocampalis
Als Gyrus cinguli wird die Windung bezeichnet, die an der Medialfläche des Großhirns über dem Balken liegt. Er wird vom Sulcus corporis callosi und Sulcus cinguli eingerahmt. Der Gyrus cinguli setzt sich vor und unter dem Balken nach vorne fort, und läßt sich hinter und unter dem Balken verfolgen (Isthmus cinguli, Gyrus parahippocampalis). Der Gyrus cinguli enthält ein Bündel von Assoziationsfasern, das vom Stirnhirn bis zur Hippocampusformation reicht.
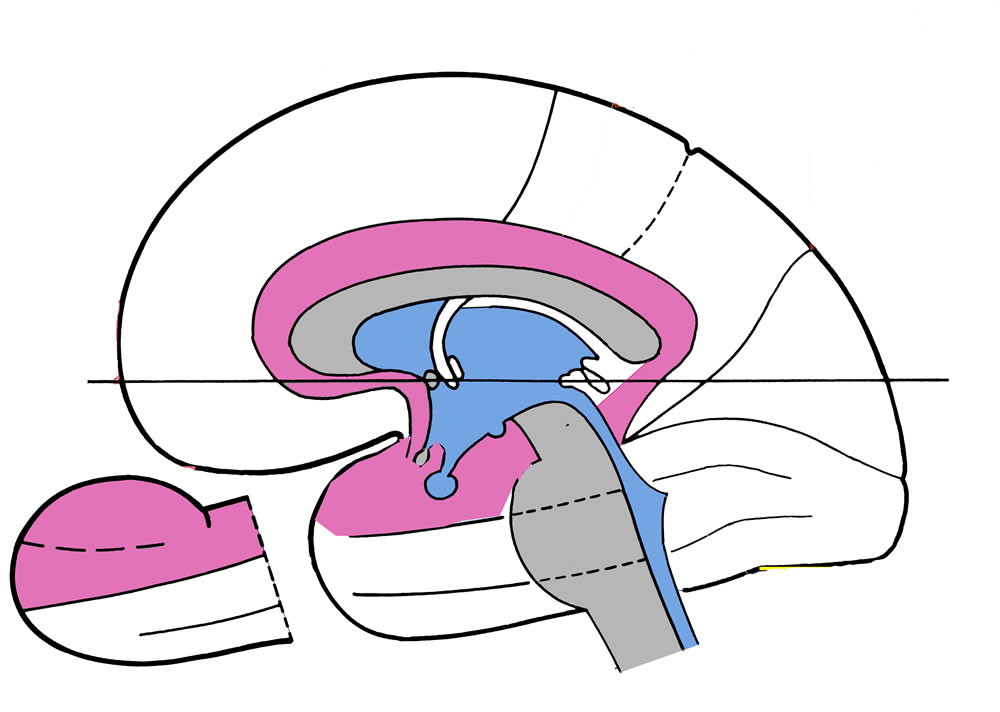
Legende
violett: (Peri-)Archicortex
blau: Ventrikel
2.2 Archicortex: Hippocampusformation
(Ammonshorn, Gyrus dentatus, Subiculum)
2.2.1. Entwicklung der Hippocampusformation
Die Entwicklung des Hippocampus läßt sich formal damit erklären, daß das progressive Wachstum des Neocortex die medial gelegenen Endhirnteile gegen den Hirnstamm drängt, so daß sich diese Windungen mangels Ausdehnungsmöglichkeiten nach innen entwickeln (Abb. 14-3).

Diese Entwicklung erklärt zwei Charakteristika der Hippocampusformation:
1. Die Rinde der Hippocampusformation wird gegen das Ventrikelsystem gedrängt. Die Fortsätze der Pyramidenzellen sind daher nicht wie im benachbarten Neocortex von weißer Substanz unterfüttert. Die markhaltigen Efferenzen von Hippocampus und Gyrus dentatus müssen sich daher zwischen Rinde und Ventrikel sammeln und bilden dort einen Markfaserüberzug (Alveus). Das Konvolut der Markfasern löst sich schließlich fransenförmig ab (Fimbria) um den Fornix (Gewölbe) zu bilden.
2. Die Einsenkung des Hippocampus und die Umstülpung des Gyrus dentatus bringt letzteren in direkte Nachbarschaft zum Subiculum und der Area entorhinalis. Fasern aus den entsprechenden Bereichen überqueren ("perforieren") den Sulcus hippocampi als Tr. perforans, bevor sie in der Hippocampusformation enden.

Legende
1, temporale Hemisphärenwand,
2, Sulcus hippocampi,
3, Gyrus dentatus,
4, Hippocampus,
5, Subiculum,
6, Cortex entorhinalis,
7, Fissura collateralis,
9, Seitenventrikel,
10, Plexus choroideus
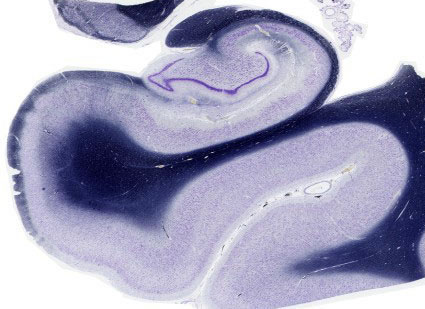
2.2.2. Struktur und Gliederung der Hippocampusformation
Als Teil des Allocortex besitzen Hippocampus und Gyrus dentatus im Vergleich zum 6-schichtigen Isocortex einen reduzierten Rindenaufbau mit nur drei Schichten (polymorphe Schicht, Pyramidenzellschicht und Molekularschicht, siehe Abb. 14-4).
Der Hippocampus besteht vorwiegend aus eng gepackten Doppelpyramidenzellen. Ihre buschigen Dendritenverzweigungen erstrecken sich sowohl in Richtung auf die piale (apikale Dendriten) wie auch auf die ventrikuläre (basale Dendriten) Oberfläche. Im Gyrus dentatus finden sich anstelle der Pyramidenzellen kleine, dicht gepackte Körnerzellen. Sie entwickeln ihre Dendritenverzweigungen in nur eine Richtung (auf die äußere Molekularschicht). Alle anderen Zellen sind Interneurone.
Abb. 14-5: Stark schematisierte Darstellung der Organisation und des Rindenaufbaus der Hippocampusformation

Abk.:
1, G. dentatus,
2, CA4,
3, CA2,
4, CA1,
5, Subiculum,
6, Cx. entorhinalis,
7, Sulcus hippocampi,
8, Sulcus collateralis,
9, Ncl. caudatus,
10, Plexus choroideus,
11, Seitenventrikel,
12, Fornix.
Beachten Sie, daß die Orientierung der Pyramidenzellen entgegengesetzt zu denen der Abb. 14-7 abgebildet ist.
Legende
Stark schematisierte Darstellung der Organisation und des Rindenaufbaus der Hippocampusformation. Die Beschreibung der inneren Organisation wird erleichtert, wenn man die Einsenkungsbewegung während der Entwicklung rückgängig macht. Die Rinde ist auf drei Schichten reduziert (polymorphe Schicht, Pyramidenzellschicht und Molekularschicht). Im Hippocampus werden sie als Stratum (Str. ) oriens, Str. pyramidale und Str. radiatum bezeichnet, im Gyrus dentatus als polymorphe Schicht (Hilus), Körnerschicht (Str. granulosum) und Molekularschicht (Str. moleculare). Die Molekularschicht des Gyrus dentatus geht in die des Hippocampus über. Aufgrund weiterer zytoarchitektonischer Merkmale werden in der Hippocampusformation die Felder CA 1 - CA 4 (für Cornu Ammonis) unterschieden. Die Zellage im Hippocampus ist zwischen 3 - 10 Zellen tief.

2.2.3 Faserverbindungen der Hippocampusformation
1. Afferenzen aus dem Cortex entorhinalis erreichen den Hippocampus über zwei Verbindungen: den Tr. alveolaris und den Tr. perforans (Abb. 14-7).
2. Afferenzen aus dem Ncl. diagonalis (=Area septi medialis) erreichen den Hippocampus über den Fornix (und Gyrus cinguli).
3. Kommissurenverbindungen vermitteln einen Faseraustausch mit dem kontralateralen Hippocampus. Diese Fasern verlaufen über die Commissura hippocampi (fornicis), die als (dünne) Faserplatte unter dem hinteren Teil des Balkens liegt.
4. Die Efferenzen des Hippocampus verlaufen über den Fornix. Sie erreichen (neben dem vorderen Kern des Thalamus und der Area septi) vor allem das Corpus mamillare. Von dort ziehen Axone über Thalamus (Ncl. anterior), Gyrus cinguli, Gyrus parahippocampalis (mit Regio entorhinalis) zum Hippocampus zurück (Abb. 14-7).

Legende
a, Afferenzen der Hippocampusformation. Cortico-hippocampales System: Vom Cx. entorhinalis (punktiert) gelangen die Informationen über zwei Faserzüge zum Hippocampus: Tr. alveolaris und Tr. perforans. Septo-hippocampale Verbindung (S): Die Fasern aus dem medialen Septumkern (Ncl. diagonalis) erreichen den Hippocampus über den Fornix.
Die Efferenzen der Hippocampusformation verlaufen über den Fornix (Fx). Er verbindet die Hippocampusformation mit dem Corpus mamillare. Von dort gelangen Fasern nach Umschaltung im vorderen Thalamuskern wieder zur Hippocampusformation zurück = Innerer Schaltkreis des Limbischen Systems (Papez-Kreis).
Abk.: AC: vordere Commissur, AS: limbische Vorderhirnregion, cm: corpus mamillare, th.a: Ncl. anterior thalami, C: Cingulum, M: Tr. mamillo-thalamicus, CC: Corpus callosum.
Der Informationsfluß im Hippocampuskomplex wird hauptsächlich über Eingänge aus dem Cortex entorhinalis gespeist (offener Pfeil). Er geht von dort streng in einer Richtung, durchläuft die verschiedenen Sektoren aber nicht stereotyp nacheinander, sondern kann einzelne Sektoren überspringen (hier dargestellt am Beispiel des Tr. perforans.
2.2.4 Schaltschema der Hippocampusformation
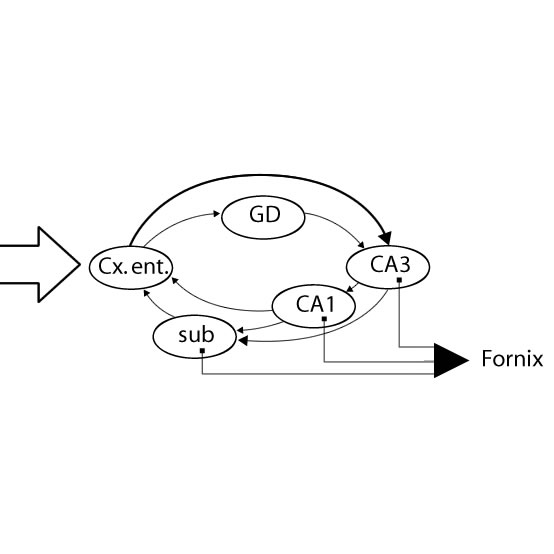
1. Das dreigliedrige (trisynaptische), offene Systems der Hippocampusformation (Tr. perforans): Fasern aus dem Cx. entorhinalis erreichen zunächst die Körnerzellen im Gyrus dentatus. Dessen Axone bilden das sog. Moosfasersystem, das an den proximalen Dendriten der Pyramidenzellen der CA 3 - Region endet. Deren Axone verlassen den Hippocampus über den Fornix. Bevor sie den Hippocampus verlassen, geben sie Axonkollaterale (Schaffer-Kollaterale) ab, die auf Dendriten der CA 1- Pyramidenzellen enden. Der Ausgang aus den CA 1- Zellen läuft dann über den Alveus ebenfalls zum Fornix und über Kollaterale zum Subiculum. Vereinfacht ergibt sich aus dieser Anordnung eine Kettenreaktion, bei der die Information nach lokaler Modulation (durch andere Eingänge und die inhibitorischen Interneurone) sukzessiv an benachbarte Hippocampus-Sektoren weitergeschoben wird. Auf diese Weise entsteht lokal eine hoch organisierte und verdichtete Stereotypie (Abb. 14-7).
Abb. 14-8: Schaltschema innerhalb der Hippocampusformation

1. Fasern aus dem Tr. perforans (TP) besitzen synaptischen Kontakt mit den Körnerzellen des Gyrus dentatus (GD).
2. Die Körnerzellen senden ihre Axone (Moosfasern, MF) zu den Pyramidenzellen der Region CA 3.
3. Letztere senden ihre Axone zum Fornix (Fx); Kollaterale dieser Axone (Schafferkollaterale, Sch-K) enden an den Pyramidenzellen in der CA 1 -Region. Die Zellen sind über 3 Synapsen (Syn. = trisynaptische Verbindung) kettenartig miteinander verbunden.
Legende
Stark vereinfachtes Modell der strukturellen Organisation im offenen (trisynaptische Verbindung) System der Hippocampusformation.
Über die Kollateralverbindungen von CA1 über das Subiculum (SUB) zum Cx. entorhinalis wird die Verschaltungskette zum Ausgangspunkt zurückgeführt (geschlossenen System der Hippocampusformation).
Beachten Sie, daß die Orientierung der Pyramidenzellen entgegengesetzt zu der in
Abb. 14-4 ist.
2. Erweiterung des dreigliedrigen Systems zur geschlossenen Schleife: Das dreigliedrige System der Verschaltung innerhalb des Hippocampus selbst ist als offenes System formuliert und läßt die intimen Beziehungen zu Nachbarstrukturen außer Betracht. Wenn man diese berücksichtigt, entsteht ein geschlossenes System in Form einer Schleife, über die die hippocampale Aktivität kontrolliert werden kann. Die Axone der CA 1 - Pyramidenzellen projezieren nämlich direkt und indirekt (über das Subiculum) zu Neuronen im Cx. entorhinalis, dem Ursprungsort des Tr. perforans und Tr. alveolaris, zurück.
2.5 Funktion der Hippocampusformation
1. Beziehung zum Kortex:
Die Hippocampusformation besitzt einerseits Verbindungen zu den Regionen, die für unsere sinnliche Wahrnehmung wichtig sind (Kortex), andererseits unterhält sie enge Beziehungen zu Gehirnteilen (Hypothalamus), die für unser Innenleben und für das Gefühl der Körperlichkeit verantwortlich sind. Es ist daher spekuliert worden, daß die Hippocampusformation Zusammenhänge zwischen inneren und äußeren Reizen herstellt und somit die Vorraussetzung für ein Gedächtnis schafft, das in einer bedeutungsvollen Beziehung zum "Ich" steht ("Ichgefühl").
2. Rolle für gewisse Aspekte von Lernen und Gedächtnis:
Es ist lange bekannt, daß der Hippocampus eine führende Rolle bei der Verarbeitung von komplexen räumlichen und zeitlichen Mustern einnimmt, da Läsionen im Bereich des Hippocampus und seiner Verbindungen (z.B. zum Fornix und Corpus mamillare) zu entsprechenden Störungen führen. In der Hippocampusformation werden Umweltinformationen nach ihrer Bedeutung für das Individuum ausgewertet. Bei entsprechender Relevanz werden diese Informationen dem Kortex zugeleitet und den eigenen Speicherressorts zugewiesen.
Besonders gut (am Zell- und Tiermodell) untersucht ist die Rolle der Hippocampusformation bei der Lern- und Gedächtnisfunktion am Beispiel der Langzeitpotenzierung (LTP) (siehe Lehrbücher der Physiologie)
3. Subkortikale Bestandteile
Hierzu gehören neben dem Mandelkern mehrere Regionen im Vorder-, Zwischen- und Mittelhirn.
Tabelle 14-2: Subkortikale Bestandteile (Subpallium) des Limbischen Systems
| Mandelkern | |
| Limbische Vorderhirnregion | (Area septi, Ncl. interstitialis der Stria terminalis, Ncl. accumbens) |
| Limbische Areale im Zwischenhirn | (Thalamus, Hypothalamus, Ncl. habenularis) |
| Limbische Mittelhirnregion | (Ncl. interpeduncularis, Area tegmenti ventralis, Ncl. tegmenti dorsalis) |

3.1 Mandelkern
3.1.1.Anatomie und Topographie
Der Mandelkern (Amygdala) liegt vor dem Unterhorn des Seitenventrikels im vorderen dorso-medialen Teil des Temporallappens. Er wird vom Allocortex (periamygdalare Rinde) bedeckt und grenzt an den Uncus des Hippocampus. Im kaudalen Abschnitt lagert sich ihm der Schwanz des Ncl. caudatus an. Der Mandelkern selbst gliedert sich in einen kortikalen und einen subkortikalen Hauptkern und den kleinen Ncl. centralis.

Legende
Grün dargestellt sind neben dem Mandelkern, der Bettkern der Stria terminalis, die Stria terminalis und die ventrale amygdalofugale Verbindung
gelb: Striatum
rot: globus pallidus, pars lat.
orange: globus pallidus, pars med.
grün: Mandelkern, der Bettkern der Stria terminalis, die Stria terminalis und die ventrale amygdalofugale Verbindung
hellblau: Thalamus
klarblau: substantia nigra, pars compacta
violett: Hippocampus, Fornix
rosa: Claustrum

Der kortikale (oder kortiko-medialer) Hauptkern ist beim Menschen schlecht entwickelt. Sein Fasersystem wird durch die Stria terminalis repräsentiert.
Der subkortikale (oder basolateraler) Hauptkern ist (beim Menschen) der größte und bestdifferenzierte Teil. Er umfaßt einen ventralen, medialen, intermediären und lateralen Unterkern. Letzterer grenzt an die ventralen Ausläufer des Claustrum. Die efferenten Verbindungen dieses Abschnittes werden durch die ventrale Mandelkernstrahlung repräsentiert.
Der Nucleus centralis zeichnet sich durch seine Hirnstammverbindungen aus.
3.1.2. Verbindungen
A, Afferenzen:
- zum kortikalen (kortiko-medialen) Kernbereich:
1. Stria olfactoria lat.: olfaktorische Fasern.
2. mediales Vorderhirnbündel: Fasern aus den limbischen Zwischenhirnabschnitten (Ncl. thalamicus med. und Ncl. ventro-med. hypothalami).
- zum zentralen Mandelkern:
3. Hirnstammverbindung: Fasern aus der Formatio reticularis des Hirnstammes (dopaminerge Neurone).
- zum subkortikalen (baso-lateralen) Kern:
4. kortikale Afferenzen: Fasern aus unterschiedlichen Kortexregionen, bes. dem temporalen
Cortex: cortex (prä-) piriformis, cortex orbitofrontalis, Gyrus temporalis inferior - visuell assoziertes Areal; Gyrus temporalis superior - auditiv assoziertes Areal.
5. Afferenzen aus dem Thalamus (Abb. 14-13).

B, Efferenzen:
Efferenzen aus dem kortikalen Kernbereich:
1. Stria terminalis: Fasern zur Area septi, zur gesamten mittleren Längszone des Hypothalamus, sowie zum Ncl. habenulae.
Efferenzen aus dem zentralen Mandelkern:
2. zu den viszeralen Hirnstammkernen.
Efferenzen aus dem subkortikalen Mandelkern:
3. zur basalen Vorderhirnregion (Stria terminalis-"Bett" = Ncl. interstitialis der Stria terminalis, Ncl. basalis)
4. Ventrale Mandelkernstrahlung: Ncl. interstitialis der Stria terminalis, Substantia innominata, seitl. Hypothalamus, Area septi, ventrales Striatum, Cortex.

3.1.3 Funktion
1. Beziehung zwischen Reiz und Affekt:
Der Mandelkern ist in den Prozeß involviert, durch den sensorische Reize motivationale und emotionale "Tönung" (Bedeutung) gewinnen. Der Wahrnehmungsprozeß ist (über Verbindungen zum Thalamus) mit Kategorisierungen verbunden, die durch Instinkt, Erfahrung oder Assoziation zu aversiven (Furcht- und Fluchttendenz fördernden) oder positiven Verhaltensweisen führen.
2. Beziehung zum Affektverhalten:
Es scheint, daß die grundlegenden Emotionen Wut, Furcht und Schrecken, die mit dem Überleben des Organismus zu tun haben, an dieses System gebunden sind. Der Mandelkern ist offenbar an der Gestaltung des affektiven Gesamtverhaltens in Bezug auf die Selbst- und Arterhaltung beteiligt und wird daher als das morphologische Substrat für das Affektverhalten betrachtet.
3. somatische Antworten:
Über die Verbindungen zum Thalamus (intralaminare Kerne, Ncl. dorsomedialis, Ncl. anterior) beeinflußt der Mandelkern die thalamo-kortikale Projektion.
4. Beziehung zum zu vegetativen Organfunktionen:
Auf der engen Beziehung zum Hypothalamus beruhen Einflüsse auf viszerale Prozesse, z.B. Nahrungsaufnahme (Eß- und Trinkverhalten, gastro-intestinale Funktion), Sexualverhalten (Zerstörungen im Bereich des Mandelkerns führen zu zwanghaften oralen und sexuellen Verhaltensäußerungen) und kardio-vaskuläre Funktion.
5. Endokrine Funktionen
6. Beziehung zum Immunsystem
3.2 Area septi
Anatomie und Topographie:
Die Septalkerne liegen beim Menschen direkt unterhalb des Septum pellucidum direkt neben der Kreuzungsstelle der vorderen Kommissur. Sie werden in eine Area septi lateralis und medialis unterteilt . Die Area septi med. ist gleichbedeutend mit dem Kern des diagonalen Bandes. Gemeinsam mit dem Ncl. basalis (Meynert) bildet er die magnozelluläre cholinerge Neuronengruppe des Endhirns (Kap. 21).
Faserverbindungen:
Die Septalregion unterhält (als einzige Region) mit allen Strukturen des Limbischen Systems rückläufige Verbindungen. Dies unterstreicht ihre Rolle als zentrale Koordinationsstelle ("großer Kommunikator"). Zusätzlich bildet sie ein Konvergenzzentrum für alle Impulse, die sich auf den Hypothalamus und seine Funktionen auswirken. Schließlich unterhält sie Verbindungen zum unteren Hirnstamm (über die Stria medullaris/Tr. habenulo-interpeduncularis sowie über das mediale Vorderhirnbündel).

Legende
Über die Area septi bestehen Beziehungen zum limbischen System, zum extrapyramidal-motorischen und sensiblen Systemen. Über den Ncl. habenulae (hab) ist die limbische Vorderhirnregion unter Umgehung des Hypothalamus mit der limbischen Mittelhirnregion (LMR) verbunden. cm, Cp. mammillare, hab, Ncl. habenulae, H, Hypophyse, CO, Chiasma opticum
Funktion:
1. Die Area septi wird für rhythmische, niedrigfrequente (4 - 7 Hertz) Wellen im Elektroenzephalogramm mit hoher Amplitude (Theta-Rhythmus) verantwortlich gemacht. Dieser Rhythmus kann in den Pyramidenzellen des Hippocampus bei bestimmten Verhaltenszuständen (Orientierungsreaktion) registriert werden.
2. Über das mediale Vorderhirnbündel übt das Septum inhibitorische Effekte auf den lateralen Hypothalamus und die Formatio reticularis aus. Tiere mit Septalläsion sind hyperaktiv.
3. Eine Stimulation der seitlichen Septalregion führt beim Menschen zu euphorischen (z.T. orgiastischen) Zuständen. Diese Zustände werden möglicherweise über die Aktivierung von dopaminergen Fasern vermittelt.
3.3 Ncl. interstitialis der Stria terminalis (sog. Bett-Kern der Stria terminalis)
3.4 Ncl. accumbens (septi) (= ventrales Striatum)
Die Anschmiegung (accumbere, lat., anschmiegen) des ventralen und vorderen Teils des Striatums an die Septalregion hat diesem ventralen Abschnitt des Striatums seinen Namen gegeben. Er gehört strukturell zum Striatum (siehe dort), ist jedoch mit dem Limbischen System funktionell sehr eng verbunden und wird als der motorische Effektorkern des Limbischen Systems betrachtet. Als Teil des Striatums verfügt der Ncl. accumbens prinzipiell über dieselben Faserquellen. Funktionell wichtig sind seine Verbindungen zur dopaminergen Area tegmenti ventralis. Der (funktionelle) Überschuß von Dopamin (an den Accumbenszellen) wird für Affektstörungen mitverantwortlich gemacht. Andererseits soll der Mangel an Dopamin für den Verlust an Emotionalität bei M. Parkinson verantwortlich sein.
3.5 Nucleus habenulae
Der Ncl. habenulae ist Teil des Epithalamus und liegt über und knapp vor der Zirbeldrüse (Abb. 12-8). Er erhält Fasern über die Stria medullaris thalami aus der Septalregion. Über die Commissura habenulae tauscht er Fasern mit dem Kern der Gegenseite aus. Aus dem Ncl. habenulae entspringen cholinerge Fasern zum Ncl. interpeduncularis, ein unpaarer Kern am First der Fossa interpeduncularis. Dieser Kern projeziert (a) zum Ncl. tegmenti dorsalis und (b) zur Formatio reticularis des Hirnstammes. Der Verbindung zwischen Area septi, Ncl. habenulae und Ncl. interpeduncularis kommt deswegen eine besondere funktionelle Bedeutung zu, weil Informationen aus dem limbischen Vorderhirn unter Umgehung des Hypothalamus an die viscerale Verteilungsstelle (Ncl. tegmenti dorsalis) sowie an die Formatio reticularis geleitet werden können (Abb. 14-9).
4. Funktion des Limbischen Systems
Die Einzelfunktionen des Limbischen Systems sind aufgrund des hohen Vernetzungsgrades seiner Instanzen schwer zu erfassen. Lokale Reizungen oder Läsionen innerhalb der einzelnen Strukturen des Limbischen Systems führen in der Regel zu multiplen Effekten, die sich oft auch an anderen Stellen auslösen lassen (z.B. Interaktion zwischen Mandelkern und Hypothalamus). Andererseits ist dieselbe limbische Funktion (z.B. ein bestimmter emotionaler oder motivationaler Zustand) durch Aktivitätsänderung in ganz verschiedenen, oft weit voneinander entfernten Teilen des Limbischen Systems auslösbar. Schließlich wirken sich verschiedene Anteile des Limbischen Systems konkurrierend oder gegensinnig auf Verhaltensäußerungen aus. Viele Befunde, die am Menschen erhoben wurden, basieren auf Stimulationsexperimenten an Patienten, bei denen bereits Störungen der Temporallappenfunktion vorlagen, oder auf Läsionsexperimenten, bei denen Verhaltenssymptome nach einer (im ethischen Sinn fragwürdigen) selektiven Zerstörung limbischer Strukturen beim Menschen registriert wurden. Verhaltenssymptome, die durch derartige Manipulationen "gebessert" wurden, betrafen Destruktivität, Feindseeligkeit und Agressivität gegenüber anderen Menschen. Einige Aspekte, die mit Funktionen des Limbischen Systems in Zusammenhang gebracht werden, sind in folgender Zeichnung zusammengefaßt:




