Das Vestibularsystem enthält Mechanorezeptoren, die Stellung und Bewegung des Kopfes im Raum registrieren.
1. Lage des Vestibularsystems
Der Vestibularapparat ist als Teil des Innenohrs beidseits im Felsenbein des Innenohrs untergebracht (Kap. 18). Die Rezeptoren befinden sich in zwei Organen des vestibulären Abschnitts des Innenohrs: den Dreh-Sinnesorganen (Bogengänge) und den Schwere-Sinnesorganen (Statolithen-Organe).

Legende
Lage des Innenohrs und Orientierung des Bogengangsystems. Blick auf die Schädelbasis. Es gibt 3 Bogengänge auf jeder Seite des Kopfes: den vorderen, hinteren und horizontalen. Die 3 Gänge liegen in 3 unterschiedlichen Ebenen, die jeweils rechtwinklig zueinander stehen. Der vordere Bogengang steht senkrecht zur Achse des Felsenbeins (1), der hintere (2) hierzu senkrecht, während der laterale (3) annähernd horizontal in der Ebene der Schädelbasis liegt. Hinsichtlich der Symmetrieachse des Kopfes sind diese Gänge paarweise so angelegt, dass jeweils ein Paar mit einem anderen zusammenarbeitet. Die horizontalen Bogengänge liegen ungefähr in der gleichen Ebene auf beiden Seiten und arbeiten hinsichtlich der Bewegungswahrnehmung zusammen. Der vordere Gang liegt ungefähr in der gleichen Ebene wie der hintere Gang der gegenüberliegenden Seite, so dass vorderer und hinterer Gang beider Seiten ebenfalls funktionelle Paarungen darstellen.


Legende
Die Ebenen der Bogengänge entsprechen nicht den Körperachsen: Vorderer und hinterer Bogengang weichen von der Median- und Frontalebene um 45 Grad ab, der seitliche, horizontale Bogengang ist nach hinten und unten um 30 Grad gegen die Horizontalebene gekippt; beide horizontale Bogengänge liegen in derselben Ebene.
c,d, Der hintere Bogengang steht senkrecht wenn der Kopf um 60° nach hinten aus der Vertikalebene gebeugt wird. 1, vorderer, vertikaler Bogengang, 2, horizontaler Bogengang, 3, hinterer Bogengang, 4, Ampulle des seitlichen Bogengangs
2. Bestandteile und Aufbau des Vestibularorgans
Das vestibuläre Labyrinth besteht aus dem Bogengangsystem (Canales semicirculares) und den Otolithenorganen (Utrikulus und Sacculus). Es ist von Perilymphe umgeben und von Endolymphe gefüllt. Die Endolymphräume sind durch (enge!) Gänge miteinander verbunden (Abb. 18-3). Jeder Bogengang besitzt an seinem Utrikulus-nahen Ende eine Eweiterung (Ampulle). In ihr befindet sich jeweils eine Epithelverdickung (Crista ampullaris) mit spezialisierten Rezeptorzellen (vestibulare Haarzellen) und Rezeptorendigungen (periphere Fortsätze von bipolaren Nervenzellen). Die Crista wird durch eine Gallertmasse bedeckt (Cupula), die ihr wie auf einem Sattel aufsitzt. Da sie bis zum Dach der Ampulle reicht, dichtet sie den Endolymphraum praktisch ab. Wenn der Kopf bewegt wird, gerät nach dem Prinzip der Trägheit die Endolymphe desjenigen Bogengangs, der in der Ebene der Bewegung liegt, in Bewegung. Als Folge kommt es zur Verbiegung der Cupula.
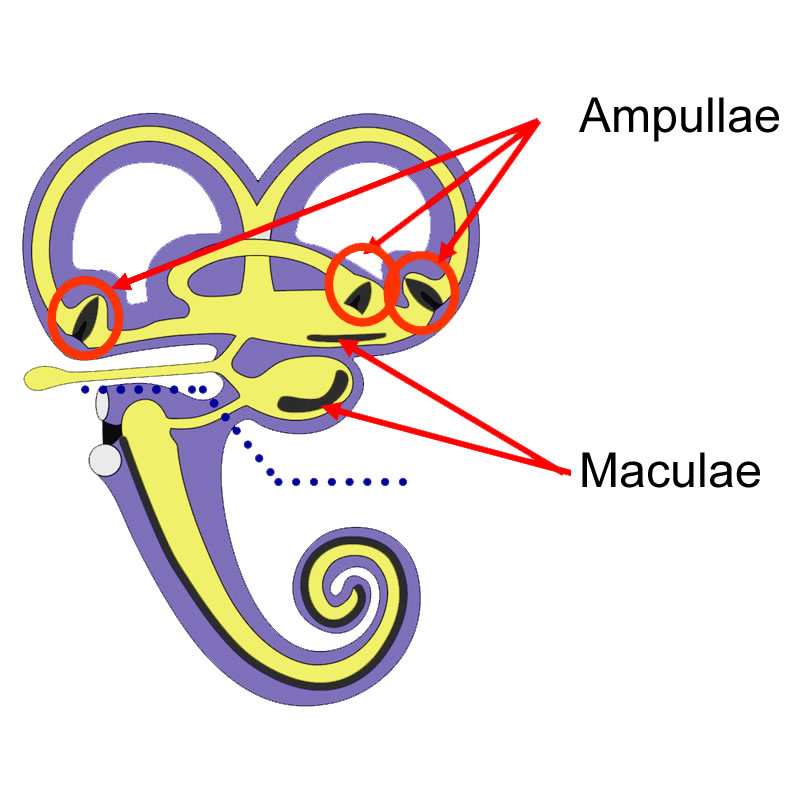

Legende
Abb. 19-4:
a, b, Sinnesepithelien des Vestibularapparats: Cristae (der Ampullen, a) Maculae (der Otolithen, b). Der Aufbau beider Sinnesepithelien ist prinzipiell gleich. Unterschiede finden sich hinsichtlich des Aufbaues der bedeckenden Gallertschicht: Sie wird bei den Cristae durch die Cupula, bei den Maculae durch die mit kristallinen Partikeln besetzte Gallertmasse (Otoconia) gebildet.
Bei den Otolithenorganen (Utrikulus und Sacculus) ist wie in den Ampullen der Bogengänge ein Abschnitt am Boden aufgetrieben. Dort befinden sich die Rezeptorzonen, die als Macula utriculi bzw. sacculi bezeichnet werden. Auch die Maculae sind von einer gallertigen Substanz bedeckt (Otoconia). In sie sind Schwerekörper eingelagert (Kalziumkarbonatkristalle, Otolithen). Die Maculae stehen zueinander annähernd senkrecht, daher reagieren sie auf unterschiedliche Reizkomponenten. Die Macula des Utrikulus liegt ungefähr in der Horizontalebene, wenn man den Kopf aufrecht hält, die Macula des Sacculus ungefähr in der Vertikalebene. Wenn der Kopf geneigt oder Linearbeschleunigungen ausgesetzt wird (z.B. Fahrt im Aufzug), wird die gallertige Masse durch die Otolithen deformiert. Dies ist mit einer Ablenkung der Haare der Rezeptorzellen verbunden (siehe unten).
3. Struktur der Rezeptorzellen (Haarzellen)
Auf der freien Oberfläche jeder Sinneszelle befinden sich ungefähr 40 bis 70 Stereozilien (stereo, lat. fest), durch parallel verbundene Aktinfilamente versteifte Mikrovilli. Zusätzlich findet sich hier ein einzelnes Kinozilium (kinein, griech. bewegen). Die Länge der Stereozilien steigt in Richtung auf das Kinozilium; daher besitzt jede Haarzelle eine morphologische Polarität (Polaritätsachse). Diese ist deswegen wichtig, weil die Haarzellen in einer sehr geordneten Art und Weise arrangiert sind und Haarzellen auf eine mechanische Lageverschiebung (Ablenkung) der apikalen Haare in einer richtungsabhängigen Art und Weise reagieren. In den Bogengängen strecken diese "Haare" in die darübergelegene Cupula, in den Maculae reichen die Kinozilien in die beschwerte Gallertmasse (Abb. 19-3).
4. Arbeitsweise
Unter Ruhebedingungen sind die Haarzellen nicht stumm, sondern zeigen eine spontane Entladungsfrequenz. Werden die Haare in Richtung auf das Kinozilium abgelenkt, führt dies zu einer Depolarisation der Haarzelle und damit zu einem Anstieg der Entladungsfrequenz der afferenten Vestibularfasern des 8. Hirnnerven. Andererseits führt eine Ablenkung der Härchen von dem Kinozilium weg zu einer Hyperpolarisation der Haarzelle und zu einer verminderten Entladungsfrequenz.
4.1 Arbeitsweise der Bogengänge
Bei einer Kopfdrehung führt die Trägheit der Flüssigkeit in den Bogengängen dazu, daß diese Flüssigkeit gegen die Cupula gedrückt wird und eine Winkelverschiebung der Sinneshaare der Rezeptorzellen auslöst. Diese Verlagerung der Cupula führt zu Verschiebungen des Rezeptorpotentials der Haarzellen der Crista und ändert schließlich die Aktivität des 8. Hirnnerven, der diese Zellen innerviert (Abb. 19-4).

Legende
Blick von oben auf die beiden horizontalen Bogengänge; sie sind an ihrer Verbindung mit dem Utriculus abgetrennt. Neben die Ampullen ist beidseits eine Haarzelle gezeichnet. Ihre morphologische Polaritätsachse weist in beiden horizontalen Bogengängen in Richtung der kürzesten Strecke zum Übergang zwischen den Bogengängen und dem Utriculus.
Abk.:
FLM: Fc. longitudinalis med. Vm,
Vs: Ncl. vestibularis med. et superior,
2: Ggl. vestibulare
RL: M. rectus lateralis,
RM: M. rectus medialis, 3, Ncll. vestibulares, (nach Kandel et al., ).
Bei einer Kopfdrehung zur linken Seite bleibt die Flüssigkeit in den Bogengängen hinter der Kopfbewegung (infolge der Trägheit) zurück. Als eine Folge wird die Flüssigkeit im linken Gang die Haarbündel in der Richtung ihrer Polaritätsachse umbiegen, während die Flüssigkeit im rechten Gang die Bündel entgegen ihrer Achse verbiegt. Die Haarzellen der linken Ampulle werden deswegen depolarisiert und Neurotransmitter freisetzen, und damit die afferenten Nervenfasern erregen. Die Haarzellen der rechten Ampulle werden jedoch hyperpolarisiert, die Entladungsfrequenz der afferenten Fasern, die diese versorgt, wird absinken. Das Gehirn wird infolgedessen 2 Zeichen dieser Kopfbewegung registrieren, einen Anstieg der Entladungsfrequenz im 8. Hirnnerv auf der einen und ein Absinken auf der anderen Seite.
Um die Arbeitsweise des Bogengangssystems zu verstehen, muß man berücksichtigen, daß die Kanäle des Bogengangssystems paarweise arbeiten. Jede Kopfbewegung führt zu einer Beeinflussung auf beiden Seiten und damit zu einer bidirektionalen Haarzellenantwort. Dem ZNS werden daher korrelierte, aber unterschiedliche Informationen aus beiden Labyrinthen zugeleitet. Die Verrechnung dieser unterschiedlichen Eingänge bildet die Vorraussetzung für die Bestimmung von Kopfbewegung und Position.
4.2 Arbeitsweise der Otolithenorgane
Die Antwort einer einzelnen makulären Haarzelle auf die Gravitationskraft, die sich auf die Otolithen und Otolithenmembran auswirkt, findet sich in Abb. 19-5. Wenn der Kopf in Horizontallage gehalten wird, wirkt sich die Gravitationskraft direkt auf die Spitze der Haarbüschel aus. Wenn der Kopf aber auf die linke Seite bewegt wird, dann werden die Haarzellen entsprechend der Achse der Polarisation abgelenkt und dies führt zu einer Depolarisation und Exzitation ihrer afferenten Fasern. Eine Bewegung auf die rechte Seite hat den entgegengesetzten Effekt. Die Folge davon ist, daß die afferente Faser, die eine individuelle makuläre Haarzelle innerviert, bei einer bestimmten Bewegung des Kopfes entweder erregt oder gehemmt wird. (Diese Beschreibung wird durch ein erstaunliches strukturelles Phänomen der Macula kompliziert: die Achsen der Haarzellen weisen nämlich nicht alle in eine Richtung; vielmehr sind sie zu einer einzelnen Kurvatur hin orientiert, die als Striola bezeichnet wird. Haarzellen im Utrikulus sind also gegen die Striola hin polarisiert. Eine Bewegung in eine bestimmte Richtung depolarisiert demnach einige Haarzellen und hyperpolarisiert andere. Dieses duale Signal hilft offensichtlich dem Gehirn, die Kopfposition mit hoher Sicherheit zu bestimmen.)


Legende
a, Kippung des Maculaorgans,
b, Aktivitätsmuster der vestibulären Haarzellen und Entladungsverhalten der afferenten Nervenfasern (N. vestibularis). Während des Ruhezustandes zeigen alle vestibulären Nervenfasern eine niedrige Entladungsfrequenz. Wenn die Haarzellen in einer Richtung (auf das Kinozilium zu) abgelenkt werden, wird diese Rate vermehrt, wenn sie in die andere Richtung abgelenkt werden, erniedrigt. Die Nervenfasern der Otolithenorgane beantworten daher eine veränderte Kopfhaltung mit einem Anstieg oder Abfall der Entladungsfrequenz.
5. Verbindungen
Die afferenten Fasern des Vestibularsystems haben ihr Soma im Ganglion vestibulare. Die zentralwärts verlaufenden Axone erreichen 4 Vestibulariskerne. Diese werden entsprechend ihrer Lage im Hirnstamm als Ncl. vestibularis lateralis, medialis, superior und inferior (oder absteigender Vestibularkern) bezeichnet. Jeder Kern besitzt individuelle Verbindungen sowohl mit dem Vestibularapparat als auch mit dem ZNS (Rückenmark, Augenmuskelkerne und Kleinhirn).

Der Ncl. vestibularis lateralis erhält Fasern aus der Makula des Utriculus sowie vom Kleinhirn (hemmende Afferenzen) und Rückenmark. Er projiziert über den Tr. vestibulo-spinalis lateralis (ipsilateral) zu den Motoneuronen der Extensoren (bis zu den lumbo-sakralen Rückenmarkssegmenten). Zusammen mit den absteigenden retikulo-spinalen Fasern, wirkt er erregend auf die Alpha- und Gamma-Motoneurone, die die Antigravitätsmuskeln der Extremitäten versorgen. Die tonische Erregung der Antigravitationsmuskeln (die Extensoren des Beines und die Flexoren der Arme) ermöglicht es uns, uns entgegen der Schwerkraft in aufrechter Lage zu halten.
Ncl. vestibularis medialis und superior erhalten ihren Eingang hauptsächlich aus den Bogengängen. Der mediale Kern entläßt zum einen Fasern in den Tr. vestibulo-spinalis medialis, der bilateral in der Zervikalregion des Rückenmarks endet. Sie bilden monosynaptische Verbindungen mit Motoneuronen, die die Halsmuskeln versorgen. Dieser Trakt nimmt an der Reflexkontrolle von Halsbewegungen teil. Zum anderen ziehen Fasern aus dem medialen und oberen Vestibulariskern zum Fc. longitudinalis medialis. Dieser Fc. verläuft durch den rostralen Pons des Hirnstammes direkt unter der Mittellinie des 4. Ventrikels und verbindet die Blickbewegungszentren des Mittelhirns, den motorischen Apparat des Halsmarks und die Vestibulaskerne untereinander. Über diese Verbindungen können die Position des Kopfes und die Augenbewegungen aufeinander abgestimmt werden (vestibulo-oculomotorische Reflexkontrolle).
Der Ncl. vestibularis inferior empfängt Fasern aus den Bogengängen und aus Utrikulus und Sacculus. Wie der Ncl. vestibularis lateralis erhält auch er zusätzlich Eingänge vom Kleinhirnwurm. Die Mehrzahl seiner efferenten Fasern verläuft über die Tr. vestibulo-spinalis und Tr. vestibulo-reticularis.
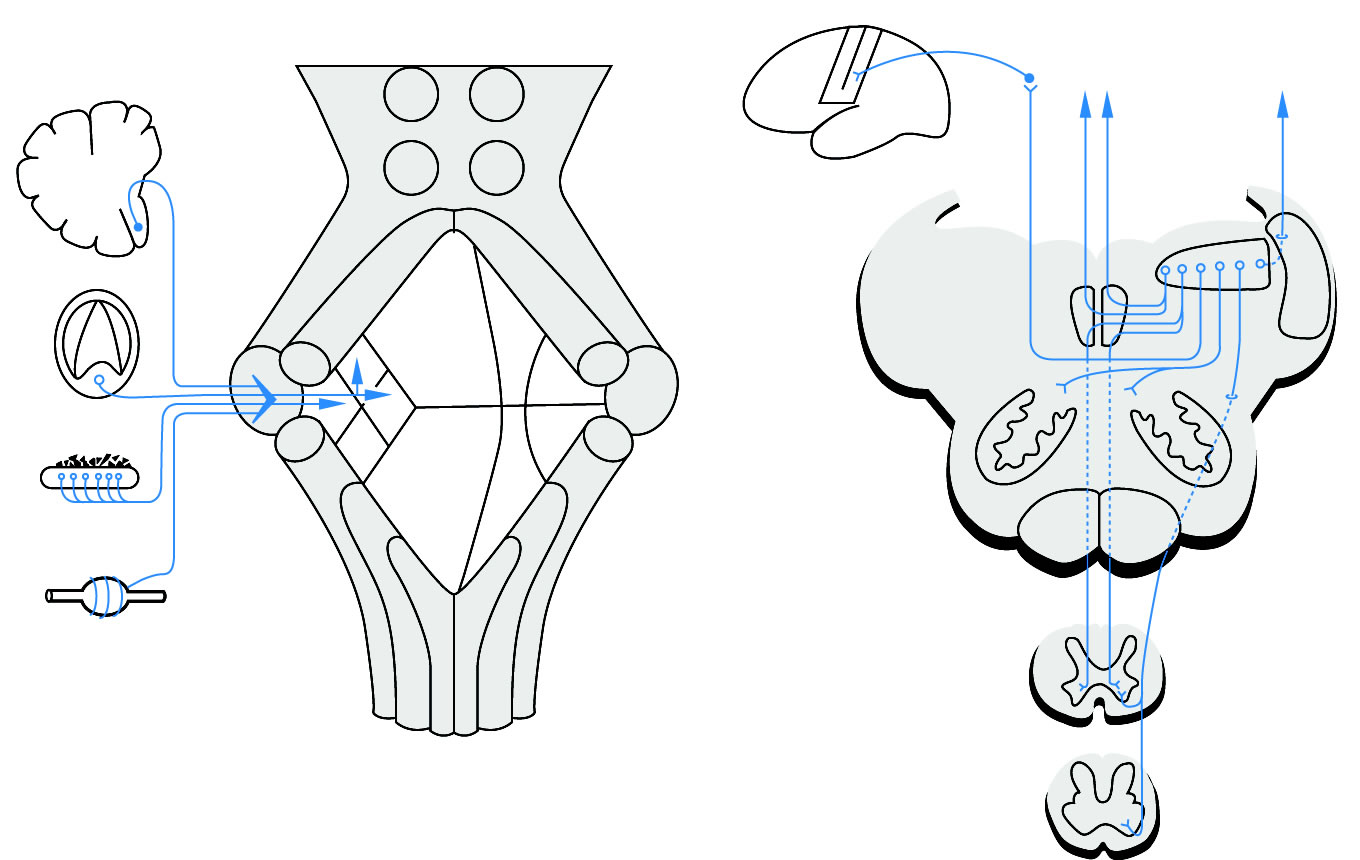
Legende
a, Afferenzen des Vestibularsystems: Informationen aus den Drehsinnesorganen (Bogengänge, dynamischer Eingang) und den Schwere-Sinnesorganen (Statolithen-Organe, statischer Eingang) werden zum vestibularen Abschnitt des Kleinhirns (dem Lobulus flocculo-nodularis) geleitet. Andere Eingänge stammen aus dem Kleinhirn (1) und aus dem Halsbereich (4). Unterschiedliche Abschnitte des Vestibularapparats sind in den Vestibulariskernen getrennt repräsentiert.
b, Efferenzen des Vestibularsystems: Die efferenten Fasern verlaufen 1. kontralateral über den Thalamus zum Kortex (zentrale Gleichgewichtsbahn), 2. ipsi- und kontralateral zum Fc. longitudinalis medialis (FLM), 3. ipsi- und kontralateral zum Tr. vestibulo-spinalis medialis, 4. ipsilateral zum Tr. vestibulo-spinalis lateralis, 5. ipsilateral zum Vestibulo-cerebellum.
Abk.: 1, Lobulus flocculo-nodularis, 2, Bogengänge, 3, Statolithenorgan, 4, Muskelspindeln, 5, Ncl. vestibularis lateralis, 6, Ncl. vestibularis superior, 7, Ncl. vestibularis inferior, 8, Ncl. vestibularis medialis, 9, Vestibularkerne, 10, unterer Kleinhirnstiel, 11, Tr. vestibulo-spinalis medialis, 12, Tr. vestibulo-spinalis lateralis, 13, Formatio reticularis, 14, Fc. longitudinalis medialis, 15, Thalamus, 16, Gyrus postcentralis.
6. Vestibuläre Reflexe
Der Vestibularapparat (Labyrinth, Vestibularkerne und zentrale Verbindungen) dient der reflektorischen Kontrolle der Körper-, Kopf- und Augenstellung bei Bewegungen, aber auch in Ruhe. Er arbeitet innig mit den Propriorezeptoren und dem optischen System zusammen. Diese funktionelle Verzahnung unterschiedlicher Sinnesinformationen (= multi-sensorische Konvergenz) erlaubt eine mehrfach gesicherte Orientierung und Bewegung im Raum, so dass der Ausfall eines dieser 3 Systeme (z.B. Gehen im Dunkeln, Zerstörung des Labyrinths) kompensiert werden kann.

Dieser funktionellen Verzahnung dienen mehrere Reflexe: Der vestibulo-spinale Reflex (VSR), der vestibulo-oculomotorische Reflex (VOR) und der optokinetische Reflex (OKN) (siehe Lehrbücher der Physiologie).
7. Funktion
Der Vestibularapparat erfüllt zwei miteinander verknüpfte Funktionsbereiche: Die dynamische Funktion, die über die Bogengänge vermittelt wird, gestattet uns Drehbewegungen des Kopfes zu erkennen und für die Reflexkontrolle der Augenbewegungen einzusetzen. Die statische Funktion, die über die Otolithenorgane (Utriculus und Sacculus) vermittelt wird, ist wichtig für die Feststellung der Position des Kopfes im Raum und spielt dadurch eine entscheidende Rolle in der Kontrolle der Haltung. Beide Teilfunktionen arbeiten zusammen, um folgende Funktionen optimal zu realisieren:
- Die Körperbalance aufrechtzuerhalten,
- koordinierte Kopf- und Körperbewegungen durchzuführen und
- die Augen auf einen Punkt fixiert zu halten, wenn das fixierte Objekt oder unser Körper bewegt wird.
8. Angewandte Anatomie
Das Vestibularsystem enthält Mechanorezeptoren, die Stellung und Bewegung des Kopfes im Raum registrieren. Normalerweise sind wir uns der vestibulären Dimension unserer Sinneswahrnehmung nicht bewusst. Sie ist jedoch für die Koordination von Augenbewegungen und motorischen Antworten sowie für die Haltung sehr wichtig. Darüber hinaus führt eine Fehlfunktion des Vestibularsystems zu Schwindel und Erbrechen - Wahrnehmungen, die uns sehr schnell bewusst werden.
8.1 Nystagmus
8.2 Schwindel
9. Zusammenfassung
Im Vestibularapparat liegen Mechanorezeptoren (vestibuläre Haarzellen) die statische und dynamische Aspekte der Vestibularisfunktion vermitteln.
Die vestibulären Haarzellen antworten auf einen Reiz entweder mit De- oder mit Hyperpolarisation in Abhängigkeit von der Richtung der Kopfbewegung. Dieses Verhalten enspricht dem der auditiven Haarzellen; es unterscheidet sich jedoch von den übrigen Mechanorezeptoren, z.B. die Paccini'schen Körperchen, die auf einen Sinnesreiz hin stets depolarisieren.
Die Vestibularapparate beider Seiten arbeiten funktionell miteinander; diese Zusammenarbeit bildet die Vorraussetzung für die Bestimmung von Kopfbewegung und Position.
Die Vestibularisfasern enden in den Vestibulariskernen und im Vestibulocerebellum. Über diese Verbindungen werden mehrere Reflexe ausgelöst. Die zentralen Verbindungen des Vestibularlabyrinths entsprechen seiner dynamischen und statischen Funktionsweise.
Die Informationen des Vestibularsystems (über Winkelbeschleunigung und Schwerkraft) werden normalerweise nicht an den Kortex weitergeleitet, d.h. bleiben normalerweise unbewusst. Obleich wir normalerweise über die Aufgaben des Vestibularsystems und seiner Leistungen nicht informiert sind, ist seine normale Tätigkeit unabdingbar für die Durchführung der meisten motorischen Prozesse.