1. Topographie
Das Kleinhirn ist eine etwa faustgroße sphärische Masse, die in der hinteren Schädelgrube liegt und vom Kleinhirnzelt dorsal abgedeckt wird. Es bedeckt die Rautengrube und ist mit Medulla oblongata und Pons durch 3 Arme (Pedunculi) verbunden.
2. Strukturelle Gliederung des Kleinhirns
Das Kleinhirn besteht aus 2 Hemisphären und einem mittleren Teil, dem Wurm (Vermis). Zahlreiche enge, querverlaufende Furchen teilen die Oberfläche in schmale, blattförmige Windungen. Diese Furchen überqueren die Mittellinie (im Gegensatz zum Großhirn) von einer zur anderen Hemisphäre ohne Unterbrechung (Abb. 9-1a). Im Sagittal-Schnittbild erinnert das Kleinhirn an eine baumartige Struktur (Abb. 9-1b). Die Windungen des Kleinhirns werden daher nicht als Gyri, sondern als Folia (Blätter) bezeichnet. Gruppen von Folia werden als Läppchen (Lobuli, L I - X) und diese wiederum zu drei Lappen (Lobi) zusammengefaßt. Der erste Lobus (L. I-V) wird als Lobus anterior bezeichnet und ist durch die Fissura prima vom 2. Lobus (L. posterior, L. VI-IX) getrennt. Der zehnte Lobulus wird als Lobus flocculo-nodularis bezeichnet, wobei der Nodulus in der Mittellinie gelegen ist während die "Flocken" beiderseits lateral liegen (Abb. 9-1a).
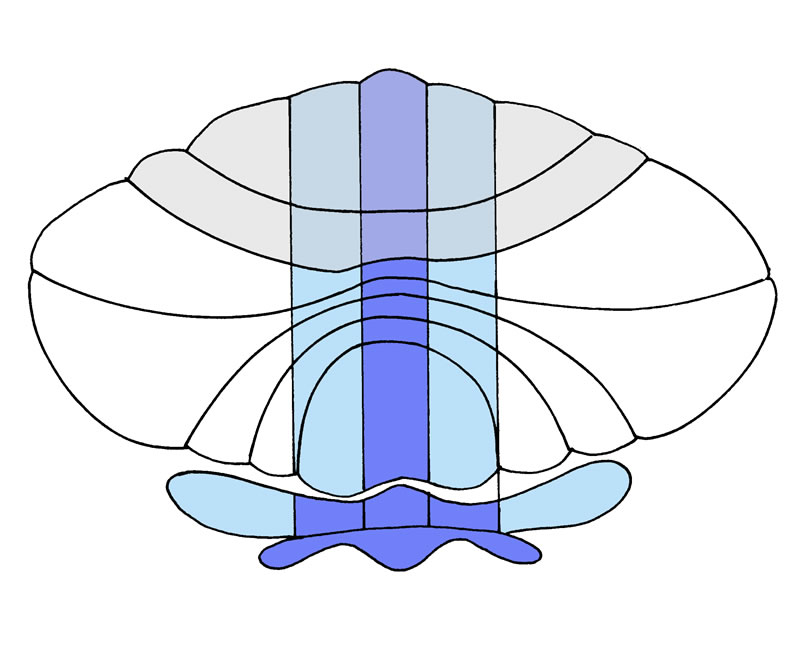

Legende
a, Gliederung des Kleinhirns in 3 Längszonen. Stark schematisierte Darstellung, bei der das gesamte Kleinhirn in einer Ebene ausgebreitet wurde (vgl. Abb. 9-1a).
Abk.: FI°, Fissura prima, FII°, Fissura posterolateralis, FL: Hemisphärenabschnitt des Lobus flocculo-nodularis.
b, Stark schematisierte Darstellung der Sagittalschnittfläche des Kleinhirns.
Die Kreissegmente teilen die drei wichtigsten Abschnitte des Kleinhirns: 1, Lobus ant., 2, Lobus posterior, 3, Pars flocculo-nodularis.
b, Ein- und Ausgänge des Kleinhirns. Sie verlaufen durch drei Paare von Faserbündeln (Pfeile). Diese werden als Kleinhirnstiele (Pedunculi cerebri) bezeichnet und bestehen aus oberem (PCS), mittleren (PCM) und unterem (PCI) Kleinhirnstiel.
Abk.: FI°, Fissura prima, FII°, Fissura posterolateralis, 1, Lobus ant., 2, Lobus posterior, 3, Lobus flocculo-nodularis, 4, Clivus, 5, Os occipitale,
3. Funktionelle Gliederung des Kleinhirns
1. Das Vestibulo-cerebellum ist mit dem Vestibularapparat verbunden
und wird durch die Pars flocculo-nodularis repräsentiert.
2. Das Spino-cerebellum erhält Afferenzen aus dem Rückenmark
und umfaßt Vermis und paravermalen Cortex (= intermediäre Hemisphäre
- Zwischenglied).
3. Das Cerebro-cerebellum erhält Eingänge aus dem Cortex cerebri
(über die Brückenkerne). Es liegt lateral zum Spino-cerebellum und umfaßt
die Hemisphären.
Diese Gliederung ist funktionell wichtig, da die 3 Zonen unterschiedliche Beiträge zur motorischen Kontrolle leisten (Abb. 9-1b).
| Kleinhirnkerne |
Ncl. fastigii |
Ncl. interpositus |
Ncl. dentatus |
| Projektionen |
Vestibulariskerne |
Ncl. ruber |
Thalamus |
|
Motorischer Kortex |
Motorischer Kortex |
Motorischer Kortex |
|
| Funktionen |
Körperbewegung |
Armbewegung |
Fingerbewegung |
| Folge von Läsionen |
Rumpfataxie |
gleichseitige Gliedmaßenataxie |
skandierende Sprache |
4. Mikroskopische Anatomie
Der mikroskopische Aufbau des Kleinhirns ist einfach. Alle 3 Teile des Kleinhirns besitzen trotz ihrer unterschiedlichen Entstehung und verschiedenen Faserverbindungen denselben histologischen Aufbau und dieselbe funktionelle Organisation. Eine dreischichtige, stark gefaltete Rinde (Cortex cerebelli) bedeckt die zentral gelegene weiße Substanz. Diese reicht in Form von Markblättern in alle Windungen. Eingebettet in die weiße Substanz liegen die Kleinhirnkerne.

Legende
Die Kleinhirnrinde besteht aus drei Schichten: Molekular-(M), Purkinje- (P) und Körnerzell- (Granular-) Schicht (G).
Abk.: MF, Moosfasern, PF, Parallelfasern, KF, Kletterfasern, A, Album, weiße Substanz. Schraffur: Pyramidenzellschicht.
4.1 Cortex cerebelli
Der Kortex enthält als charakteristische Zellelemente die Purkinjezellen (PZ). Sie stellen die einzigen Projektionsneurone des Kortex dar. Die Somata liegen in einer einzigen gleichnamigen Schicht; ober- und unterhalb hierzu liegen zwei weitere Schichten: die oberflächenzugewandte Molekularschicht, die tieferliegende Körnerzellschicht.

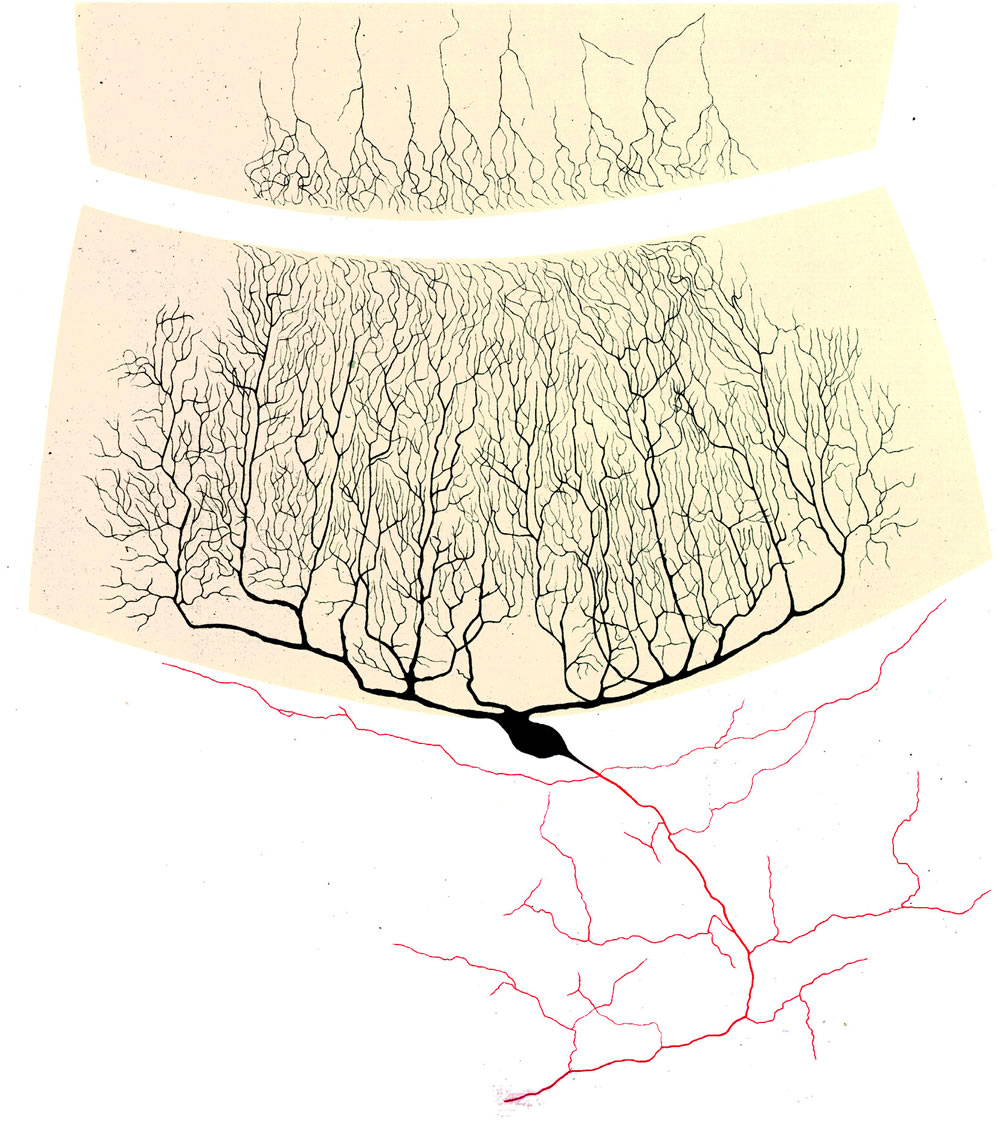
Alle anderen Zellen des Kortex gehören zur Klasse der Interneurone (Körnerzellen, Golgizellen, Stern- und Korbzellen). Die Körnerzellen (KZ) bilden die einzige Klasse exzitatorischer Neurone. (Sie können als "verlängerter Arm" der exzitatorischen Moosfasern betrachtet werden, siehe unten). Ihr dünnes aufsteigendes Axon teilt sich in der Molekularschicht unter Bildung von exakt parallelen Fasern, deren Spannweite bis zu 6 mm reichen kann (Parallelfasern, s.o.).
Die Golgizellen liegen in der Körnerschicht. Sie zeichnen sich durch ihre polare Orientierung und ihren reichen Dendritenbaum aus. Sie erhalten ihren Eingang in der Körnerschicht über Kletter- und Moosfasern und in der Molekularschicht aus den Parallelfasern. Ihr Axon endet nach reger Verzweigung in sog. Glomerula der Körnerschicht.
Die Stern- und Korbzellen liegen in der Molekularschicht. Sie erhalten ihre Eingänge durch die Parallelfasern. Ihre Fortsätze sind wie die Dendriten der PZ quer zur Längsachse der Folia und damit senkrecht zu den Parallelfasern orientiert. Der Name Korbzellen besagt, daß ihre Axone die PZ umwickeln.
Mit Ausnahme der Körnerzellen sind alle Zellen des Cortex cerebelli inhibitorisch: die PZ, die Korb- und Sternzellen, sowie die Golgizellen.
4.2 Eingänge in den Cortex cerebelli
Die Kleinhirnafferenzen enden direkt oder indirekt an den PZ. Sie werden durch zwei erregende Fasersysteme gebildet: durch die Kletterfasern und das Moosfasersystem (Abb. 9-4a).
Die Kletterfasern (Abb. 9-3b) stammen aus der unteren Olive. Ihre Axone kreuzen die Mittellinie, verlaufen durch den unteren Kleinhirnstiel und enden am Dendritenbaum der PZ. Jede PZ unterhält nur mit einer Kletterfaser Beziehung (allerdings kann sich jede Kletterfaser mehrmals teilen). Die PZ ist damit in Hinsicht auf die Kletterfasern "monogam", die Beziehung ist "fokusiert". Zu den Kleinhirnkernen geben die Kletterfasern jeweils eine Kollateralfaser ab.
Das Moosfasersystem (Abb. 9-3b) setzt sich aus Moosfasern, Körnerzellen und Parallelfasern zusammen. Die Moosfasern stammen aus vielen Quellen (Vestibularkerne, Rückenmark, Formatio reticularis, Kortex). Auch sie geben eine Kollateralfaser zu den Kleinhirnkernen ab. Die Moosfasern teilen sich intensiv bevor sie an den Körnerzellen enden. Das Axon der Körnerzellen steigt durch die PZ-Schicht in die Molekularschicht auf, teilt sich dort in 2 Äste, die wie der Horizontalbalken eines "T" exakt entgegengesetzte Richtungen einschlagen. Diese Fasern verlaufen streng parallel zueinander; daher haben diese Fasern den Namen "Parallelfasern" erhalten. Die Richtung dieser Parallelfasern entspricht der Ausrichtung eines Foliums und steht damit senkrecht zur Ebene der PZ, so daß deren Dendritenbäume (durch bis zu 20 000 Parallelfasern) senkrecht gekreuzt werden (Abb. 9-2d). Der Impuls der Moosfasern setzt sich daher über die Körnerzellen zu den PZ fort (daher Moosfaser-"System"). Die Unterbrechung mit starker lokaler Verzweigung in der Körnerzellschicht erhöht die Zahl der PZ, die letztlich durch ein einzelnes Moosfaser -Axon stimuliert werden. Wegen der starken Verzweigung der Moosfasern und der enormen Länge der Parallelfasern von bis zu 6 mm ist der Impulsstrom über Moosfasern, Körnerzellen (KZ) und Parallelfasern nicht fokusiert.
Moosfasern besitzen keine direkte Verbindung mit den PZ. Die Unterbrechung des Moosfasersystems in der KZ-Schicht führt zu Divergenz und Flexibilität zu Lasten der Örtlichkeitsauflösung.



Legende
a, Grundprinzip der Verschaltung im Kleinhirn. In der primären Verschaltungsebene (I) werden die Kleinhirnkerne (KK) erregt. Die sekundäre Ebene (II) führt entweder zur Erregung (IIa) oder (über inhibitorische Interneurone, IN) zur Hemmung (IIb) der Purkinjezellen (PZ). Die PZ üben selbst eine Hemmung auf die Kleinhirnkerne aus. Damit treten Aktivierungs- und Hemmungselemente auf der Ebene der Kleinhirnkerne in Wettstreit.
b, Kletterfaser-Verbindungen
c, Moosfaser-Verbindungen
Der einzige Ausgang des Cortex cerebelli erfolgt über die PZ und ist inhibitorisch.
4.3 Kleinhirnkerne
Die PZ-Axone verlaufen durch die Körnerschicht und die weiße Substanz zu den Kleinhirnkernen, wo sie inhibitorisch wirken. Jeder Kleinhirnkern enthält PZ-Zustrom aus dem Kleinhirnsegment, das direkt über ihm steht. Dem Vermis sind der Dachkern (Ncl. fastigii) und Teile der Vestibulariskerne, dem intermediären Rindenteil ist der Ncl. interpositus (Ncl. emboliformis und Ncl. globosus), den Hemisphären der Ncl. dentatus zugeordnet. Die Kleinhirnkerne sind alles andere als reine Durchgangs- oder Verteilerstationen. Vielmehr bestimmt das synaptische Geschehen an dieser Stelle wesentlich die Kleinhirnfunktion.
5. Schaltschema
Das elementare Verschaltungsmuster beruht darauf, daß die beiden Eingänge (Kletter- und Moosfasern) einerseits die Kleinhirnkerne, andererseits die PZ erregen. Da die PZ hemmend auf die Kleinhirnkerne wirken, steht die Interaktion der erregenden Einflüsse (durch das Eingangssignal) und der hemmenden Einflüsse (durch das im Kortex bearbeitete Signal) in den Kleinhirnkernen im Vordergrund (Abb. 9-5a). Um sich das Zusammenwirken dieser beiden Elemente zu veranschaulichen, sollte man zunächst die Einzelkomponenten getrennt betrachten.


5.1 Kletterfaser - Verbindung:
Die Aktivierung der Kletterfasern führt zu einer lokal begrenzten, sehr kräftigen Erregung der PZ. Gleichzeitig werden Golgizellen aktiviert, die den Moosfaser-Eingang hemmen. Beides, die kräftige Aktivierung der PZ und die Blockierung der Moosfaser-Afferenzen, bedeutet, daß sich die Kletterfaser-Eingänge bei ihrer Aktivierung der PZ voll bemächtigen und konkurrierende Eingänge seitens der Moosfasern ausschalten (Abb. 9-4b,c).
5.2 Moosfaser - Verbindung
Die Erregung von Moosfasern aktiviert über die Körnerzellen und Parallelfasern einen ganzen Stapel von PZ-Dendriten; zusätzlich werden die 3 Typen inhibitorischer Interneurone erregt, die den Reiz räumlich und zeitlich begrenzen (Abb. 9-4b,c): die Korb-, Sternzellen, und die Golgizellen. Das Gesamtresultat dieser Interaktion führt dazu, daß nach einer kurzen Entladungssalve einer relativ großen, aber sehr gut definierten PZ-Population das Aktivierungsmuster blitzschnell zusammenbricht.
5.3 Die Beziehung zwischen Kleinhirnkortex und Kleinhirnkernen
Sowohl die Kletter- wie die Moosfaserafferenzen führen zur Erregung von PZ, die ihrerseits eine hemmende Wirkung auf die Kleinhirnkerne ausüben. Jede Moos- und Kletterfaser-Afferenz besitzt eine erregende Kollateral-Verbindung zu den Kleinhirnkernen. Daher führt jeder Impuls, der das Kleinhirn erreicht, zu einer direkt erregenden und verzögert hemmenden Beeinflussung des Ausgangskerns des Kleinhirns. Die Neurone der KH-Kerne erhalten daher über Kollaterale eine Direktkopie der Information, die dem Kleinhirnkortex zugeleitet wird, außerdem über die Axone der PZ eine Kopie der in der Kleinhirnrinde bearbeiteten Information. Diese Organisation reflektiert offenbar die Aufgabe des Kleinhirns, einen Vergleich der Information vorzunehmen.
6. Verbindungen des Kleinhirns
6.1 Afferenzen
Die Eingänge zum Kleinhirn stammen aus dem Rückenmark, dem Hirnstamm
und dem Cortex cerebri.
1. Die Verbindungen mit dem Rückenmark (ipsilaterale spino-cerebellare
und cuneo-cerebellare Verbindungen, siehe Kap. 10) vermitteln Rückmeldesignale,
die es dem Kleinhirn gestatten, einen Informationsabgleich zwischen der
zentraler Zielvorgabe (Programmierung der Bewegung) und der aktuellen
Bewegungslage (Propriozeption) durchzuführen.
2. Die Verbindungen mit dem Hirnstamm betreffen Kerne, die ihrerseits
Afferenzen aus dem Rückenmark erhalten und daher als indirekte
Kleinhirnafferenzen aufgefaßt werden können: Tractus spino-olivo-cerebellaris
(mit Unterbrechung in der unteren Olive) und Tractus spino-reticulo-cerebellaris
(mit Unterbrechung im N. reticularis lateralis). Außerdem treten über
den Tr. nucleo-cerebellaris propriozeptive Fasern aus den N. trigeminus
in das Kleinhirn (Kap. 10).
3. Die Informationen aus dem Cortex cerebri verlaufen (vorwiegend) über
die cortico-ponto-cerebellare Bahn. Sie stammen von Neuronen aus allen
Cortexarealen und ziehen durch den mittleren Kleinhirnstiel in die kontralaterale
Hemisphäre. Diese Verbindung dient der Programmierung und Durchführung
von Bewegungen.
6.2 Efferenzen
Das Spino-Cerebellum projeziert über die Ncll. fastigii und interpositus
zum Rückenmark. Beide Kerne beeinflussen Motoneurone über absteigende
Bahnen. Axone aus den Ncll. fastigii schließen sich Projektionen an, die
medial im Rückenmark absteigen (Tr. reticulo- und Tr. vestibulo-spinalis).
Axone aus dem Ncl. interpositus verlaufen lateral im Rückenmark (Tr. rubro-spinalis).
Außer diesen absteigenden Fasern gibt es solche, die über den seitlichen
Thalamus zum Kortex projezieren.
Das Cerebro-Cerebellum projeziert über den Ncl. dentatus kontralateral
zum einen über den ventrolateralen Thalamuskern zum motorischen Kortex,
zum anderen zum kleinzelligen Ruberkern. Von dort nimmt die rubro-olivo-cerebellare
Verbindung ihren Ursprung.
Das Vestibulo-Cerebellum projeziert zum Ncl. fastigii. Einige Axone gelangen auch direkt zu den Vestibularkernen. Diese Verbindung ist aus der Phylogenese zu verstehen (der Ncl. vestibularis lateralis kann als Bestandteil des Kleinhirns verstanden werden). Zusammengefaßt wird der gesamte Ausgang des Kleinhirns über die Kleinhirn-und Vestibularkerne geleitet.

Legende
Die Afferenzen und Efferenzen verlaufen durch die drei Kleinhirnstiele.
a, Afferenzen. 1. Information aus dem Rückenmark (Moosfasereingang) über den unteren und oberen Kleinhirnstiel. 2. Verbindungen mit dem Hirnstamm (über die untere Olive und die Formatio reticularis) über die unteren Kleinhirnstiele. 3. Eingang aus dem Cortex cerebri über die Nuclei pontis über die mittleren Kleinhirnstiele.
b, Efferenzen: Der Ausgang des Cerebro-cerebellums erfolgt über den Ncl. dentatus, und beeinflußt die motorische und prämotorische Region des Kortex. Die cerebello-spinalen Projektionen laufen über die Ncll. fastigii und interpositus und kontrollieren die medialen und lateralen absteigenden Systeme. Das Vestibulo-Cerebellum projeziert zum Ncl. fastigii und zu den Vestibularkernen.
Abk.: 1, Lobus ant., 2, Lobus posterior, 3, Lobus flocculo-nodularis, 4,Brückenkerne, 5, Oliva inf., 6, Ncl. ruber, 7, Ncl. dentatus, 8, Ncl. fastigii, 9, Vestibulariskerne, 10, Formatio reticularis.
7. Funktion
Der innere Aufbau des Kleinhirns ist stereotyp. Funktionelle Unterschiede zwischen verschiedenen Bereichen des Kleinhirns ergeben sich daher hauptsächlich aus seinen Faserverbindungen.
Eingänge aus dem Vestibulocerebellum dienen der Koordination der Motorik mit Information aus dem Vestibularapparat. Damit sichert das Vestibulocerebellum die Haltemotorik (Antigravitationsmuskulatur; über Macula sacculi und utriculi vermittelt) und die Folgemotorik der Augenbewegungen (über das Ampullensystem vermittelt).
Das Spinocerebellum sorgt über Rückenmarkverbindungen für die Koordination spinaler unwillkürlicher Bewegungsabläufe (z.B. beim Gehen und der Gestik) und der Körperstellung (Stellreflex) und zwar unter Koordination von Impulsen aus den Muskel- und Hautrezeptoren, sowie den Hirnnervenkernen.
Das Cerebrocerebellum unterstützt und kontrolliert die vom Großhirn initiierten Willkürbewegungen. Fein abgestimmte, koordinierte Präzisionsbewegungen, die durch Exaktheit, Stetigkeit und Symmetrie gekennzeichnet sind, werden durch die reflektorische Regelung des Muskeltonus seitens des Kleinhirns ermöglicht.
Über die Verbindungen mit dem Cortex cerebri und Rückenmark ist das Kleinhirn in der Lage, Abweichungen der geplanten Bewegungsabläufe noch während des Bewegungsvorganges auszugleichen (und zwar durch die Abstimmung externer und interner Feedbacksignale), um dadurch Bewegungsentwurf und tatsächliche Bewegungsdurchführung optimal anzupassen. Die Aufgabe des Kleinhirns liegt daher in der indirekten Unterstützung von Haltung und Bewegung, indem es die Tätigkeit verschiedener motorischer Zentren miteinander koordiniert und dabei besonders die zielgerechte Durchführung der vom Großhirn entworfenen Willkürbewegungen und deren Koordination mit motorischen Aktivitäten (Tonus, Haltung, Gleichgewicht) gewährleistet. Die Aufgabe des Kleinhirns läßt sich daher auch als reflektorisches Koordinationssystem oder als ein von sensiblen Einflüssen geregeltes Rückmeldesystem beschreiben.
8. Angewandte Anatomie
Das Fehlen einer direkten Beziehung zur sensiblen (Ausnahme: Vestibularapparat) und motorischen Funktion impliziert, daß eine Läsion oder eine Zerstörung des Kleinhirns weder zu sensiblen Ausfällen noch zur Lähmung führt. Trotzdem sind Läsionen im Kleinhirn durch gut charakterisierte und oft schwerwiegende Beeinträchtigungen gekennzeichnet, die dazu führen können, daß sogar einfachste motorische Sequenzen nicht begonnen werden können bzw. die motorischen Zielbewegungen ausbleiben oder fehlläufig sind. Die Symptome von Kleinhirnstörungen lassen sich meist auf folgende 2 Grundstörungen zurückführen:
8.1 Die zerebellare Ataxie:
Störungen, bei denen Bewegungskoordination. gestört ist.
8.2 Muskelhypotonie
Ermüdungserscheinungen der Muskeln (Asthenie).
Prinzipiell führt die Kleinhirnsymptomatik aus folgenden Gründen zu ipsilateralen
motorischen Symptomen:
1. Die (meisten) Kleinhirnafferenzen verlaufen ipsilateral.
2. Kleinhirnefferenzen (über den Thalamus zum motorischen Kortex) verlaufen gekreuzt. Da die absteigenden motorischen Verbindungen (z.B. Pyramidenbahn, Tr. rubro-spinalis, Tr. rubro-bulbaris) ebenfalls gekreuzt verlaufen, stellt sich der Verlust motorischer Koordination ebenfalls ipsilateral dar.
9. Zusammenfassung
Das Kleinhirn besteht aus 2 Hemisphären und einem Wurm (Vermis).
Das Kleinhirn wird durch zwei tiefe Querfurchen in drei Lappen geteilt.
Es gibt drei Kleinhirnabschnitte mit unterschiedlicher Zuordnung zum
Vestibularapparat (Vestibulo-Cerebellum), Rückenmark (Spino-Cerebellum)
und Kortex (Cerebro-Cerebellum).
Die Kleinhirnrinde (Cortex cerebelli) besteht aus drei Schichten: Molekularschicht,
Purkinjezellschicht und Körnerzellschicht. Zentralwärts schließt sich
das Marklager an.
Die morphologisch auffallendsten Zellen des Kleinhirns sind die Purkinje-Zellen
(PZ). Sie werden durch zwei Fasersysteme erregt: durch die Kletterfasern
und das Moosfasersystem.
Die Kletterfasern stammen aus der unteren Olive. Sie enden am Dendritenbaum
der PZ. Zu den Kleinhirnkernen geben die Kletterfasern jeweils eine
Kollateralfaser ab.
Das Moosfasersystem setzt sich aus Moosfasern, Körnerzellen und Parallelfasern
zusammen. Die Moosfasern stammen aus vielen Quellen. Die Moosfasern
selbst besitzen keine direkte Verbindung mit den PZ.
Die PZ-Axone enden an den Kleinhirnkernen, wo sie inhibitorisch wirken.
(Danben gibt es eine direkte Verbindung zu den Vestibularkernen.
Das Schaltschema des Kleinhirns basiert auf der Interaktion der erregenden
Eingänge aus dem Kletterfaser- und Moosfasersystem auf der einen Seite
und dem hemmenden Eingang aus den Purkinjezellen auf der anderen.
Das Vestibulo-Cerebellum dient der Aufrechterhaltung des Gleichgewichts
und der Kontrolle der Augenbewegungen. Das Spino-Cerebellum ist wichtig
für die Kontrolle der Körpermuskulatur, das Cerebro-Cerebellum ist wichtig
für die Planung von Bewegungsabläufen.
Schädigungen des Kleinhirns machen sich ipsilateral bemerkbar.



