Das somatische NS wird in das zentrale (ZNS)
und periphere Nervensystem (PNS) geteilt. Das PNS umfaßt
die Gesamtheit aller Hirn- und Rückenmarksnerven sobald diese die Rückenmarkssegmente
oder den Hirnstamm verlassen haben, außerdem alle Ganglien und Rezeptoren,
soweit sie außerhalb des ZNS liegen. Es stellt damit das afferente und
efferente nervöse Leitungssystem dar.
Gliederung des peripheren Nervensystems.
| PNS | A. Hirnnerven (10 Paare*) | |||||
| B. Rückenmarksnerven (31 Paare) | 1. Cervicalnerven | (C) | 8 Paare | |||
| 2. Thorakalnerven | (Th) | 12 Paare | ||||
| 3. Lumbalnerven | (L) | 5 Paare | ||||
| 4. Sakralnerven | (S) | 5 Paare | ||||
| 5. Coccygealnerv | (Co) | 1 Paar | ||||
* Die Hirnnerven I und II sind Leitungsbahnen, die dem ZNS zugerechnet werden.
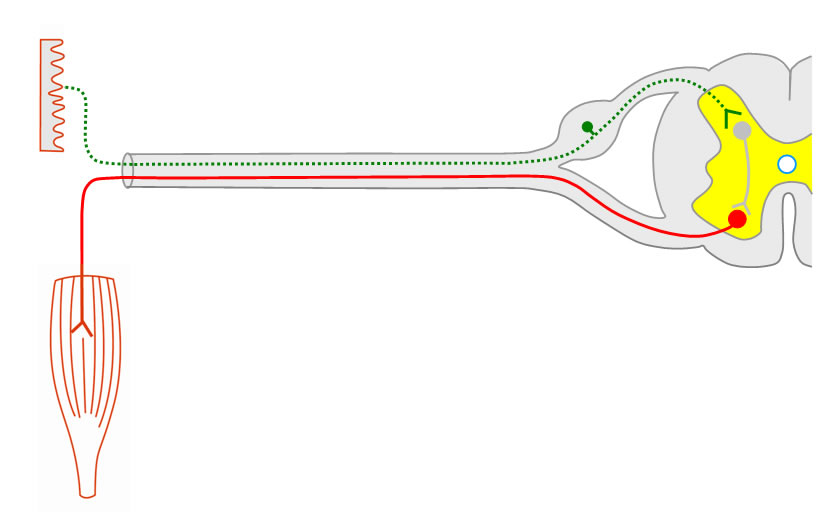
1. Rückenmarksnerven
Sensible, afferente Fasern treten über die hintere Wurzel in das Rückenmark ein; motorische, efferente Fasern treten über die vordere Wurzel aus.
Informationen, die für die Peripherie bestimmt sind (efferente Fasern),
verlaufen durch die vordere Wurzel, während Informationen aus der Peripherie
(afferente Fasern) über die hintere Wurzel in das Rückenmark eintreten.
Beide Wurzeln treffen sich und bilden einen gemischten peripheren Rückenmarksnerven.
An jeder dorsalen Wurzel der 31 Rückenmarksnerven findet sich ein Spinalganglion
kurz vor der Vereinigung mit der ventralen Wurzel zum Spinalnerven.
1.1 Peripherer Nerv
siehe auch Abb. XX Kapitel 1
Im peripheren Nervensystem verlaufen die Nervenfasern gebündelt und bilden einen makroskopisch präparierbaren (peripheren) Nerv, der durch das Epineurium vom umgebenden Gewebe abgegrenzt wird. Er besteht aus einer wechselnden Anzahl von langen Axonbündeln, die ihrerseits von einer konzentrisch geschichteten straffen bindegewebigen Hülle (Perineuralscheide, Perineurium) umgeben sind. Jedes Axonbündel enthält schließlich unterschiedlich viele parallel verlaufende (markhaltige oder marklose) Axone. Einige Axone sind Fortsätze motorischer Neurone, die zur Muskulatur ziehen; andere sind Teil sensibler Neurone, die Information in das zentrale NS leiten. Damit sind die Qualitäten eines peripheren Nervs gemischt. Leitungsbahnen des PNS zeigen erhebliche Unterschiede gegenüber denen des ZNS.
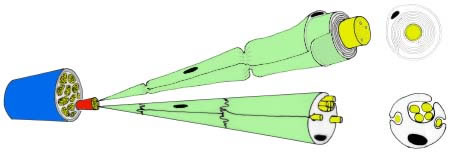
Legende
Aufbau eines peripheren Nerven mit Hervorhebung der Unterschiede zwischen markhaltigen (oben) und marklosen Axonen.
Epineurium: blau,
Perineurium: rot;
das Endoneurium liegt der Schwannzell-Membran (grün) auf.
Tabelle 4-1: Unterschiede der Organisation von weißer Substanz im peripheren und zentralen NS
| PNS | ZNS |
|---|---|
| Axone besitzen keinen knöchernen Schutz | Schutz durch Schädelkapsel oder Wirbelkörper |
| Axone verlaufen in Hirn- oder Rückenmarksnerven | Axone verlaufen in Tractus |
| Ansammlungen der Nervenzellkörper liegen in Ganglien | Ansammlungen der Nervenzellkörper liegen in Kernen |
| Nervenfaserbündel werden durch Perineurium umschlossen | Axonbündel werden von der Leptomeninx umgeben |
| Axone werden von Endoneurium und Neurolemm (Schwann'sche Scheide) umgeben | Axone werden von Neuroglia umgeben |
| Myelin stammt von Schwann-Zellen | Myelin stammt von Oligodendrozyten |
· Plexus finden sich im Bereich von Hals, oberer Brust und Lendenwirbelsäule (Plexus cervicalis, brachialis, lumbosacralis).
· In den Plexus werden Nervenwurzeln gebündelt und dann auf die verschiedenen Nerven aufgeteilt.
· Durch die Plexusbildung wird der Einfluß / das Versorgungsgebiet der einzelnen Rückenmarksegmente ausgedehnt.
Nervengeflechte (Plexus):[11]
Als Plexus wird die makroskopisch erkennbare Verflechtung von Leitungsbahnen bezeichnet. Nervenplexus sind Verbindungen der
Rami ventrales der Rückenmarksnerven (z.B. Plexus cervicalis oder brachialis). In ihnen werden die Fasern umgruppiert, so
daß jeder Rückenmarksnerv Fasern mit anderen peripheren Nerven austauscht. Als ein Resultat dieses Faseraustausches
ergibt sich, daß die motorischen und sensiblen Versorgungsgebiete der Fasern, die durch eine Wurzel verlaufen und solcher
Fasern, die in einer einzelnen peripheren Nervenfaser verlaufen, unterschiedlich sind (daher müssen Sie später die
radikuläre=segmentale von der peripheren Innervation unterscheiden können).
Da die Nerven segmental ein/austreten, und
die zugeordneten Gewebeanteile (Somite) ursprünglich auch segmental angelegt sind, wird das von einem einzelnen
Spinalnerv innervierte Gebiet als Segment bezeichnet. Die begrenzte periphere Region, die über die hintere Wurzel
innerviert wird, wird als Dermatom bezeichnet, während die entsprechende motorische Region Myotom genennt wird.
Je nachdem, ob ein sensibler, motorischer oder gemischter Nerv betroffen ist, liegen sensible und/oder motorische Ausfallserscheinungen vor. Aus der Organisation der peripheren Nerven ergibt sich, dass die Durchtrennung eines distalen Teils eines peripheren Hautnerven im Verlust der/von Oberflächensensibilität in einer umschriebenen Fläche resultiert. Dies steht im Gegensatz zur Verletzung der hinteren Wurzel eines Spinalnerven, die häufig nur zu einer relativ milden abgeschwächten sensiblen Empfindungsstörung führt. Ähnlich verhält es sich bei den motorischen Nerven: Durch die Plexusbildung wird erreicht, dass ein Muskel aus mehreren Segmenten innerviert wird. Der Ausfall eines Spinalsegmentes führt daher (noch) nicht zu Lähmung eines Muskels; sie tritt (erst) nach Durchtrennung eines peripheren Nerven ein.
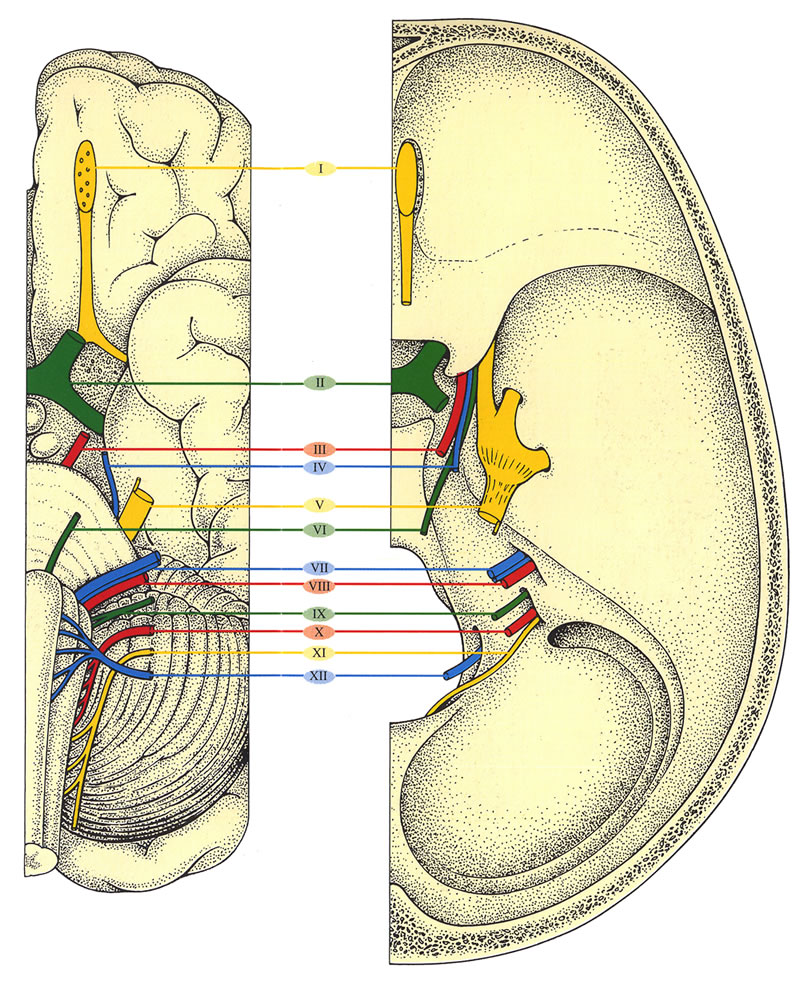
2. Hirnnerven, Nn. craniales (CN)
Die Nummerierung der 12 Hirnnervenpaare erfolgt entsprechend
ihrem Durchtritt durch die Dura mater in fronto-okzipitaler Folge. Einige
von ihnen sind nur motorisch, andere nur sensibel, der Rest ist gemischt.
Die Hirnnerven III-XII entspringen dem Hirnstamm und innervieren Kopf, Hals und spezielle (höhere) Sinnesorgane. Darüberhinaus überwindet der CN X (N. vagus) den Kopf- Halsbereich und gelangt zu den Brust- und Baucheingeweiden. Sie vermitteln die
- motorische und sensible Innervation von Kopf und Hals
- Innervation von speziellen Sinnesorganen
- parasympathische Innervation autonomer Ganglien (die Eingeweidefunktionen wie Atmung, Herzschlag, Kreislauf, Blutdruck, Husten, Schlucken usw. regulieren).
Die Organisation dieser Hirnnerven entspricht prinzipiell
denen von Rückenmarksnerven; die Existenz von speziellen Sinnesorganen
und die komplizierte Entwicklung im Kopf machen die Anatomie allerdings
unübersichtlicher.
Rückenmark:
- 2 Klassen von afferenten (sensiblen) Nervenfasern (somato-afferente und viszero-afferente Fasern) und
- 2 Klassen von efferenten (motorischen) Nervenfasern (somato-motorische und viszero-motorische = autonome) Fasern
Hirnstamm: wie Rückenmark plus:
- "spezielle" sensible = sensorische Fasern und
- "spezielle" motorische = branchiomotorische Fasern.
Dadurch gibt es im Hirnstamm 3 Arten von efferenten und
3 Arten von afferenten Neuronen.
Im Gegensatz zu den Rückenmarksnerven, die jeweils alle Klassen von Fasern enthalten,
sind einige Hirnnerven nur motorisch, andere nur sensibel, der Rest ist gemischt.
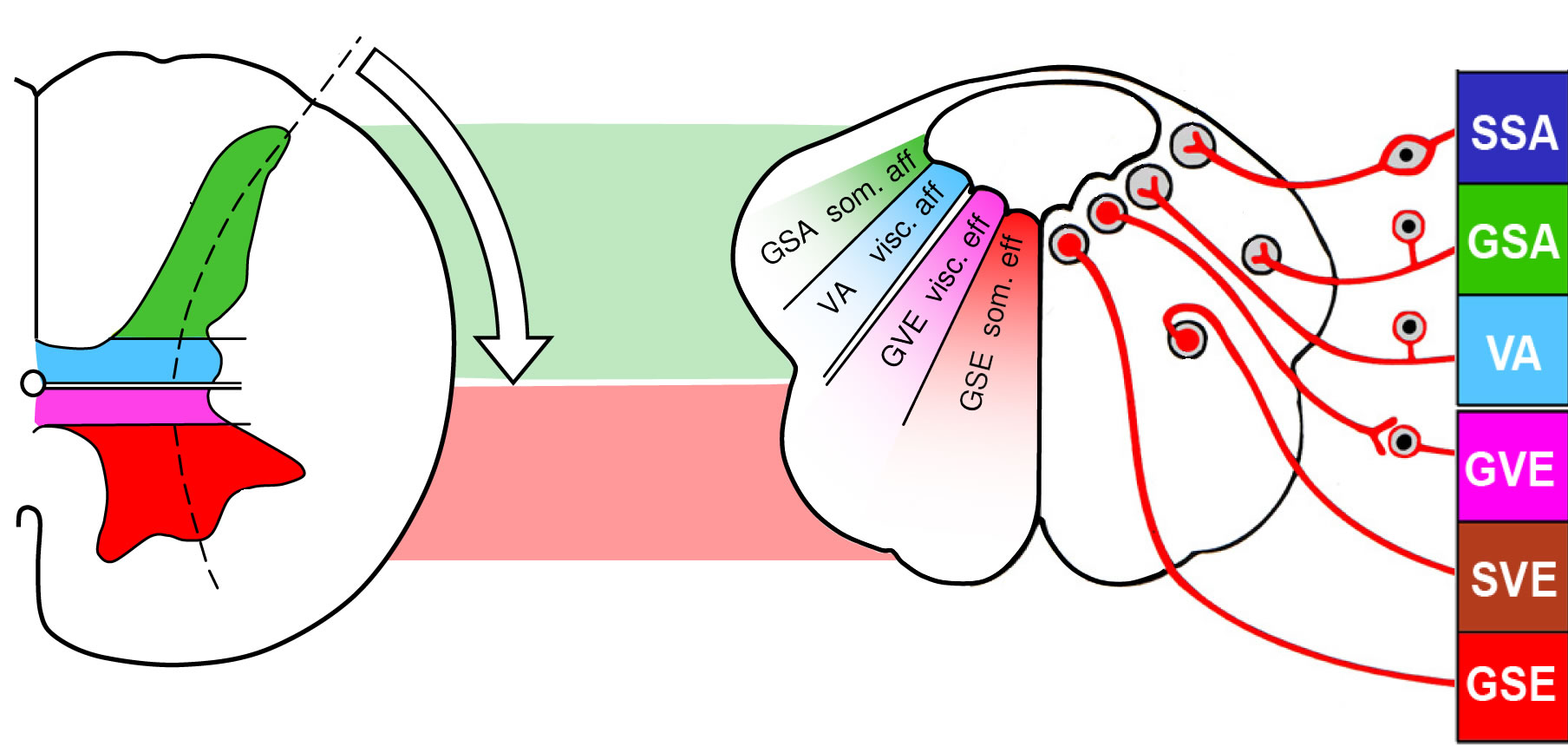
Legende
Abb. 4-4:
links: Die Rückenmarksnerven enthalten afferente (sensible) und efferente (motorische) Fasern mit jeweils
somatischer und viszeraler Qualität. Die Hirnnerven (III-XII; Hirnstamm-Nerven) enthalten ebenfalls afferente (sensible)
und efferente (motorische) Fasern mit jeweils somatischer und viszeraler Qualität. Diese dem Bauplan von Rückenmark und
Hirnstamm gemeinsamen Komponenten werden als generelle Komponenten bezeichnet. Zusätzlich werden jedoch im Hirnstamm
Strukturen innerviert, die es auf der Rückenmarksebene nicht gibt:
1. Abkömmlinge des Kiemendarms (Muskeln, die sich
nicht wie im Rückenmark aus den Myotomen ableiten, sondern aus den Eingeweiden).
2. Sinnesorgane (Innenohr und
Geschmacksorgan). Da diese Komponenten im Rückenmark nicht auftreten, werden sie als spezielle Komponenten bezeichnet, um
die Besonderheit gegenüber dem generellen Bauplan zu betonen. Damit besitzen Hirnnerven neben den 2 Klassen von
generell-afferenten (SSA, GSA) Nerven und 2 Klassen von generell-efferenten (GSE, GVE) Nerven noch jeweils
speziell-viszerale (VA) und speziell-somatische (SVE) Nerven.
3. Organisation von Sinnessystemen
Als Teil der Umwelt benötigt unser Organismus Strukturen,
die Informationen (Reizen, Energieformen) aus der Umwelt aufnehmen
und sie dem ZNS zuleiten, wo sie nach Berechnung und Bewertung für die
eigenen Lebensäußerungen und Überlebensstrategien nutzbar gemacht werden
können. Zusätzlich verarbeitet der Organismus Informationen, die seiner
Selbstorganisation dienen. Die Strukturen zur Registrierung derartiger
Reize werden als Rezeptoren bezeichnet. Sie sind in bestimmten
Regionen konzentriert und werden in ihrem Leistungsbereich meist durch
Hilfszellen beeinflußt. Die Summe der Rezeptoren, die ähnliche Informationen
(Modalitäten) vermitteln, und ihre Hilfsstrukturen werden als Sinnesorgane
bezeichnet.
3.1 Einteilung der Rezeptoren
Tabelle 4-2: Klassifizierung von Rezeptoren und der von ihnen vermittelten Empfindungen
Klasse der Rezeptoren |
vermittelte Sensibilität |
Rezeptortyp |
Qualität der Empfindung |
|---|---|---|---|
| EXTERO-REZEPTOREN | spezielle Sensibilität (Sensorik) | Lichtrezeptorzelle, Haarzelle, Geschmackszelle, Riechzelle | |
| generelle Sensibilität der Körperoberfläche
(Somato-,Haut-Sensibilität), epikritische Sensibilität, protopathische Sensibilität |
Dermato-Rezeptoren: a. Mechano-Rezeptor b. Thermo-Rezeptor c. Schmerz-Rezeptor. |
a. Druck, Berührung, Vibrationsempfindung (Pallästhesie), Kitzel b. Kälte, Wärme, Hitze c. Information über Gewebsgefährdung: Oberflächen-, Tiefenschmerz |
|
| PROPRIO-REZEPTOREN | Tiefensensibilität der Gliedmaßen: | Rezeptoren der somatischen Afferenzen: a. Muskelspindeln b. Sehnenorgane c. Mechano-R des Vestibularapparats |
Stellungs-, Bewegungsänderung (Kinästhesie = Bewegungs- oder Muskelsinn), Gleichgewichtssinn |
| VISCERO-/ = ENTERO-REZEPTOREN | Sensibilität der Eingeweide | Rezeptoren der viszeralen Afferenzen (Eingeweide- oder
viszerale R.): a. Mechano-Rezeptor b. Chemo-Rezeptor c. Osmo-Rezeptor d. Schmerz-Rezeptor |
in der Regel geht die Aktivierung der Viszerorezeptoren nicht mit einer Empfindung einher (evtl. Tiefenschmerz) |
Legende
Unter "protopathischer" Sensibilität versteht man grobe Druck-, Berührungs-, Schmerz- und Temperaturempfindungen, die als verschwommen, diffus empfunden werden und stark affektgebunden sind. Als "epikritische" Sensibilität werden Druck-, Berührungs- (diskriminatorische Wahrnehmung) und Vibrationsempfindung bezeichnet.
3.2 Eigenschaften von Rezeptoren
- a. Spezialisierung auf bestimmte Reizformen [12]
- b. Begrenzung des Leistungsumfangs [13]
- c. Qualität [14]
- d. Spezifische Sensibilität[15]
- e. Selektion des Informationsflusses. [16]
3.3 Morphologie von Rezeptoren
Nicht alle Rezeptoren depolarisieren in Antwort auf einen entsprechenden Reiz. Einige Rezeptoren, wie z. B. die Photorezeptoren, hyperpolarisieren auf den Reiz hin. Die vestibulären Haarzellen antworten entweder mit De- oder mit Hyperpolarisation in Abhängigkeit von der Richtung oder Neigung der Kopfbewegung.
Der Aufgabenbereich eines Rezeptors betrifft die Umwandlung
eines physikalisch/chemisch dimensionierten Reizes in Nervenerregung =
elektrisches Signal. Dieser Prozeß (Transduktion) findet
an der Rezeptormembran statt. Bei der Beschreibung der Rezeptormorphologie
müssen daher der Mechanismus der Umwandlung des Reizes in ein elektrisches
Signal und die strukturelle Beschreibung der Rezeptortypen unterschieden
werden.
Primäre und sekundäre Sinneszellen: Bei einer primären Sinneszelle findet die Transduktion am Rezeptorende eines sensiblen Neurons statt; bei einer sekundären Sinneszelle befindet sich zwischen Rezeptor und sensiblem Neuron eine Synapse (Sehzelle, Geschmackszelle, Haarzelle).
3.4 Oberflächen-Sensibilität
Topographie der Oberflächen-Rezeptoren
Da sich alle für ein Axon typischen Strukturen im Rezeptorende weiter verfolgen lassen, spricht man hier auch von einem Rezeptoraxon, obgleich es sich um rezeptive (ihrer Eigenschaft nach dendritische) Zellfortsätze handelt.
Oberflächen-Rezeptoren sind Endigungen sensibler Nervenfasern in der
Haut. Ihre Topographie innerhalb der behaarten und unbehaarten Haut ist
charakteristisch.
Morphologie der Oberflächen-Rezeptoren
- Freie Nervenendigung (NE)
- NE mit Hilfsstrukturen (Kapsel, Kern).
Abb. 4-5: Klassifizierung der Oberflächen-Rezeptoren
|
Endigungen |
Struktur |
Merkmale |
|
|---|---|---|---|
| unbehaarte Haut | behaarte Haut | Beispiele | |
| Nervenendigungen (NE) ohne Hilfsstrukturen | |||
| Freie NE: verzweigte und unverzweigte Nervenendigung in der Epidermis und Dermis |

|

|
Freie unverzweigte (a) und verzweigte Nervenendigung (b): Die freie NE stellt das terminale Segment eines myelinisierten oder nicht-myelinisierten Rezeptoraxons dar. Es ist von Schwann-Zellen umhüllt. Die Umhüllung ist nicht komplett; entblößte Partien des Axons (Protuberanzen), die nur noch von Basallamina bedeckt sind, treten durch Lücken zwischen den Schwann-Zellen. |
| Nervenendigungen mit Hilfsstrukturen | |||
|
Kontakt mit sekundärer Sinneszelle
|

|

|
Merkel-Zellen (c):Am Grund der Epidermiszapfen im Str.basale gelegene sekundäre Sinneszellen (modifizierte Keratinozyten). Sie stehen in Kontakt mit dem Rezeptor-Axon, das in Form einer Scheibe (Tastmeniskus) gebaut ist. Mikrovilli-artige Fühler dienen der Vermittlung der mechanischen Energie .
Die Merkel-Tastscheibe (d):bildet eine umschriebene Struktur, die aus bis zu 70 Zellen besteht. Die Achse der einzelnen Merkelzellen ist parallel zur Oberfläche orientiert, die NE liegen immer auf der dermalen Seite. |
| verzweigte NE mit Kapselapparat (Kapsel und Kern) |

|

|
Meissner-Tastkörperchen (e): Die Schwann-Zellen (Kern) sind horizontal lamelliert; zwischen ihnen liegen stark verzweigte NE. Die Kapsel wird durch unregelmäßige Lagen von Perineurium- Zellen und Kollagenfasern gebildet. Mechanischer Anschluß an das Filament-(Tonofibrillen-) System der benachbarten Epidermiszellen durch feine Kollagenfaserbündel .
Haarfollikelrezeptor (f): Die NE sind so um das Haar angeordnet, daß der sensorische Spalt radiär zum Haarschaft steht. Bei Bewegung des Haares wird auf die NE ein mechanischer Reiz ausgeübt. |
| verzweigte NE mit Kapselapparat unter Einbeziehung funktionell spezialisierter Kollagenfasern |

|
Ruffini-Körperchen (g): Ausgezogene, spindelige Organe von 1-2mm Länge. Die sehr gering lamellierte Kapsel umschließt einen flüssigkeitgefüllten Raum. Er ist von gerichteten Kollagenfasern durchzogen, die durch die Pole der Kapsel hindurch in Beziehung zu Kollagenfasern der umgebenden Dermis treten. (Vgl. Aufbau der Muskel- und Sehnenspindeln). | |
| unverzweigte, gerade NE (Lamellenkörperchen;. einfache oder komplexe eingekapselte Körperchen. > |

|
Vater-Pacini-Körperchen (h): Das Rezeptoraxon dieses sehr komplexen eingekapselten Körperchens liegt mit seinem gestreckten unverzweigten Innenkolben in einer hoch geordneten lamellierten Hülle aus Schwann-Zellen (= innere Lamellenschicht, Kern) und Perineural-Zellen (Kapsel). | |
Legende
Abk.: BL, Basallamina, Ka, Kapsel, Ke, Kern, M, Markscheide, NE, Rezeptoraxon, P, "Protuberanzen", Ausstülpungen der axoplasmatischen Membran, S, Schwann-Zelle, T, Tastmeniskus. Beachten Sie, daß die Größenunterschiede der verschiedenen Rezeptoren unberücksichtigt sind.
Das Rezeptoraxon beginnt an der Stelle, an der das Axon seine geschlossene
Schwann-Zellhülle (bzw. Markscheide) verliert. Seine Struktur ist bei
allen Dermato-Rezeptoren gleich, d.h. morphologisch gibt es kein Merkmal,
das einen bestimmten Dermato-Rezeptor auszeichnet. Die Unterschiede ergeben
sich allein durch die Modifikation der Hilfsstrukturen.
Einteilung der Oberflächenrezeptoren
- topographisch (Lagebeziehung zu den verschiedenen Schichten der Haut),
- genetisch (Beziehung zu Zellen des Ekto-, Meso-, Entoderms)
- funktionell (Struktur-/Funktionsbeziehung:
Mechanorezeptoren, Thermorezeptoren, Chemorezeptoren, Nozizeptoren (Schmerzrezeptoren).
Mechano-Rezeptoren: Struktur-/ Funktionsbeziehung
Die Zuordnung morphologisch definierter Rezeptortypen zu den verschiedenen Sinnesmodalitäten der Haut ist durch die sog.
Mikroneurographie möglich. Dabei wird die Größe der rezeptiven Felder und das Entladungsverhalten einzelner
Rezeptoren durch die Ableitung einzelner Nervenfasern unter verschiedenen Reizbedingungen registriert (Abb. 4-7).
Abb. 4-7: Anordnung zur Registrierung des Antwortverhaltens einer einzelnen afferenten Faser des N. medianus

Legende
Anordnung zur Registrierung des Antwortverhaltens einer einzelnen afferenten Faser des N. medianus, in die eine Mikroelektrode eingeführt wurde (nach D Vallbo et al.).
Nach Bestimmung der rezeptiven Felder und des adäquaten Reizes ergeben sich für die Mechanorezeptoren folgende 4 Haupttypen mechanosensibler Einheiten:
Abb. 4-8: Beziehung zwischen rezeptiver Feldgröße, Adaptationsverhalten und verschiedenen Mechanorezeptoren.
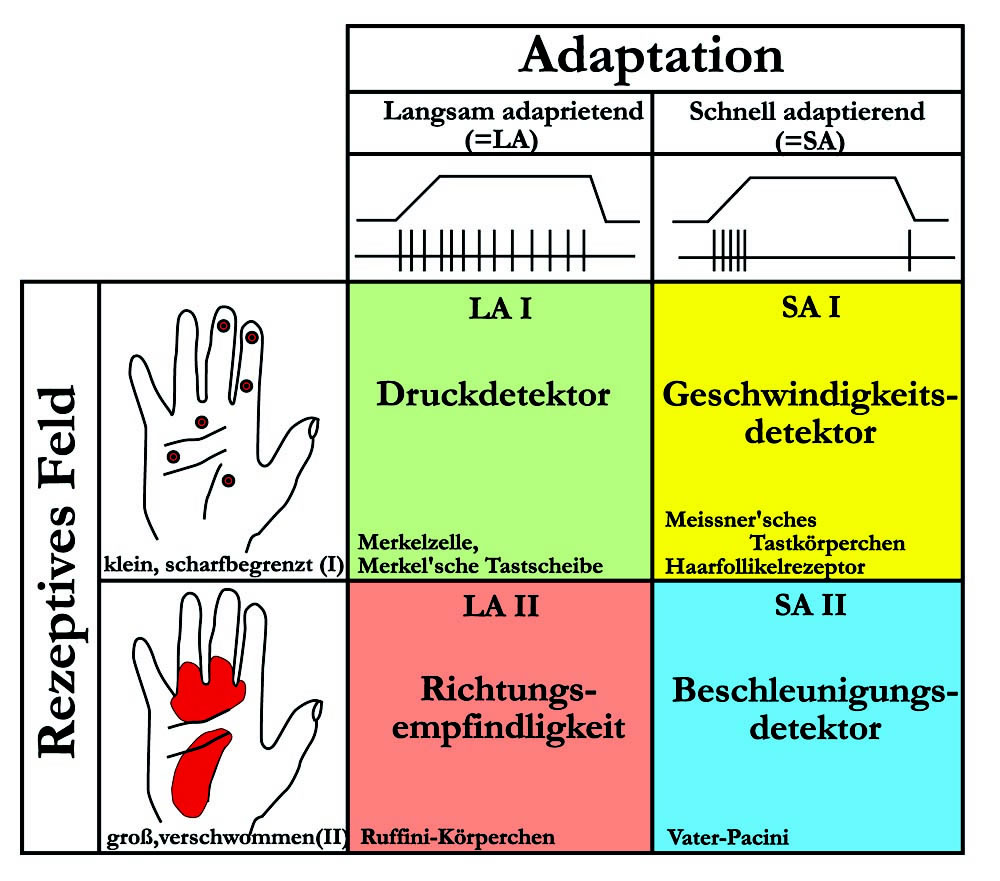
Thermorezeptoren
Wärmeempfindung wird durch Kälte- und Wärmerezeptoren vermittelt. Temperaturempfindung ist punktuell: getrennte Kälte- und
Wärmepunkte entsprechen diskreten Innervationszonen. Morphologisch handelt es sich um dünne, myelinisierte Fasern mit
Verzweigung an der Epidermis/Dermisgrenze. Thermorezeptoren besitzen kleine rezeptive Felder und ein spezifisches
Reiz/Antwortverhalten.
Chemorezeptoren
Der adäquate Reiz für Chemorezeptoren sind chemische Substanzen. Dies sind Geruchs- und Geschmacksstoffe (siehe Kap. 16 und Kap. 17), aber auch CO2, NaCl.
Nozizeptoren (Schmerzrezeptoren)
Die biologische Bedeutung der Schmerzempfindung liegt in der Warnung des Organismus vor Schädigungen seines Gewebes und
pathologischen Störungen des Zellstoffwechsels. Der adäquate Reiz für die Erregung eines Nozizeptors liegt daher in einer
Gewebeschädigung. Sie veranlaßt die Bildung und Freisetzung von Substanzen (Bradykinin, Histamin, Serotonin, Zytokinen),
die zur Aktivierung der Nozizeptoren und Erregung der nozizeptiven Afferenzen führen. Nozizeptoren können auch durch
mechanische und thermische Reize erregt werden. Die Haupttypen der Nozizeptoren heißen daher: Mechanische Nozizeptoren -
werden durch starke mechanische Reize aktiviert und zwar vorwiegend durch scharfe Objekte. Hitze-Nozizeptoren -
reagieren, wenn das Rezeptoraxon auf über 45° C aufgeheizt wird. Polymodale Nozizeptoren - reagieren gleichermaßen auf
alle Arten schädigender Reize (auf mechanische, thermische und chemische Noxen) (siehe Kap. 10).
3.5 Tiefensensibilität
· entstehen innerhalb des Körpers,
· können wahrgenommen werden (Lage- oder Kraftsinn, Kinesthesie) oder unbemerkt bleiben;
· stehen im Dienst der Koordination von Bewegung.
Die Rezeptoren der Tiefensensibilität werden Propriozeptoren genannt. Es sind "Eigen-" Rezeptoren des Halte- und Bewegungsapparates. Sie liegen in Muskeln und Sehnen und werden daher als Muskelspindeln und Sehnenorgane (Golgi-Organe) bezeichnet. Sie reagieren auf Zustandsänderungen des Bewegungsapparates (Lage und Bewegung) und sind an der Regulation der Motorik beteiligt.
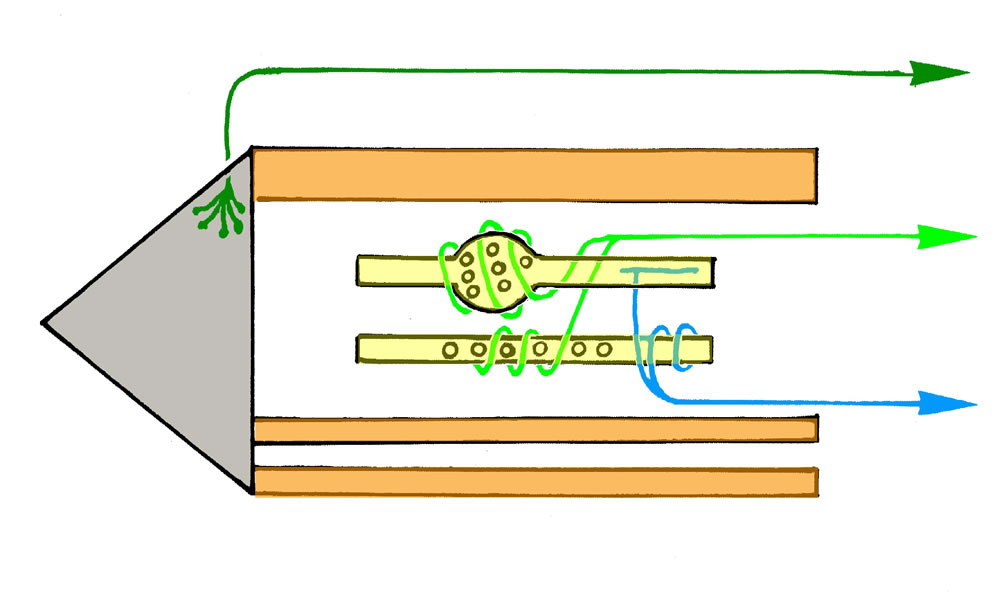
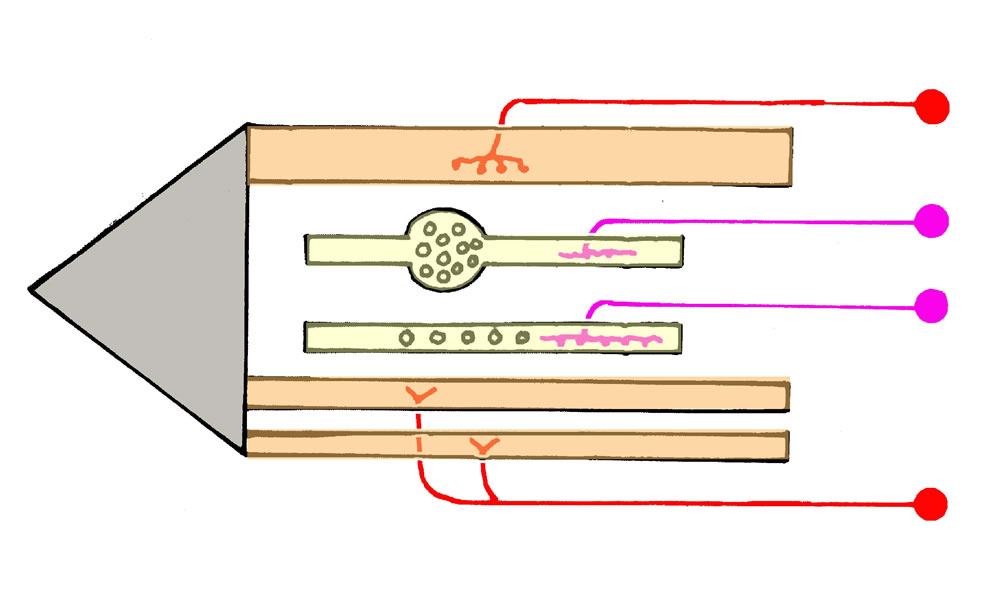
Legende
a, Sensible Endigungen im Bereich der Muskelspindeln. Ia: primär sensible Endigung; die dicke, myelinisierte Faser endet im
Äquatorialbereich der intrafusalen Muskelfaser (Anulospiralendigung). II: sekundär sensible Endigung; sie wickelt sich um
die beiden Pole der intrafusalen Muskelfaser. Ib-Fasern. Sie verzweigen sich zwischen den Sehnenfasern. Bei Dehnung der
Sehnenfasern werden sie erregt (Messung der Muskelspannung).
b, Motorische Innervation. Die Endplatten der γ-Motoneurone
(γ-MN) enden polwärts von der efferenten Innervation. Ihre Aktivierung führt zur Kontraktion der Polregion und zur
Dehnung der Zentralregion (führt zur Erregung der Ia-Afferenzen).
4. Weiterleitung der Sinnesreize zum Rückenmark
Die Zellkörper der sensiblen Nervenfasern liegen in den Spinal- und Hirnstammganglien (pseudounipolare Ganglienzellen). Ihre proximalen Fortsätze treten über die hintere Wurzel in das Rückenmark ein (siehe dort).
5. Efferenzen des NS
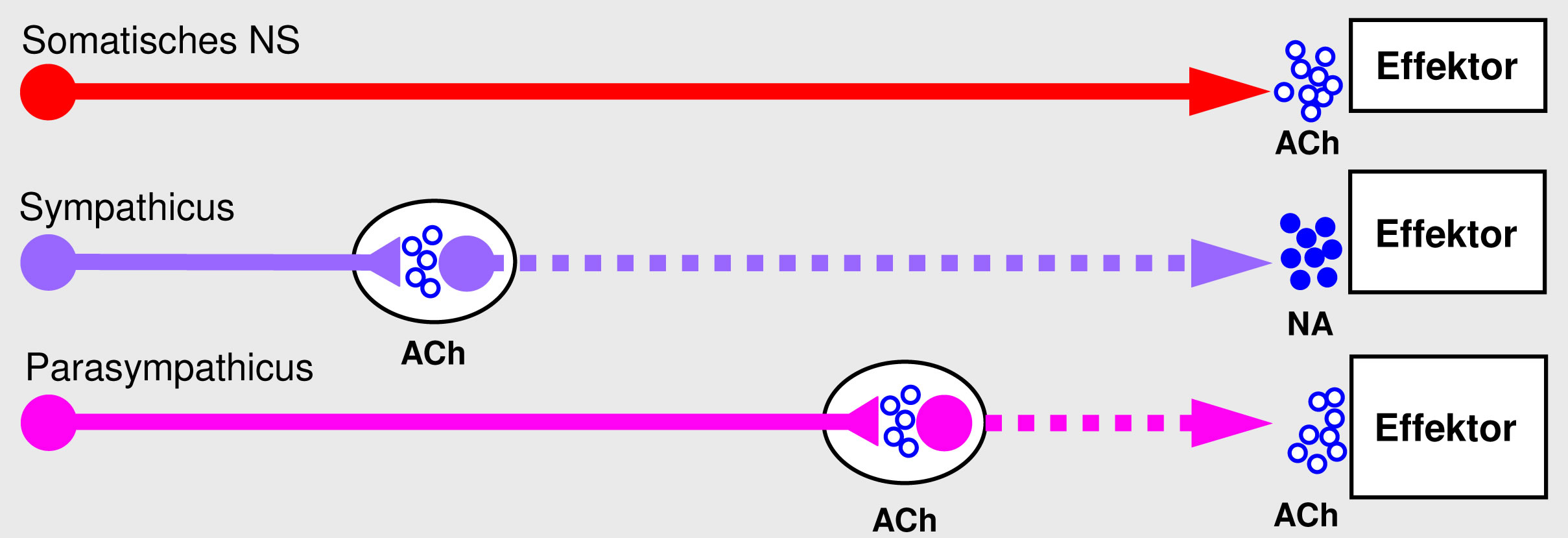
Legende
Abb. 4-11
Alle Axone, deren Soma im ZNS liegt (die somatischen und präganglionären Fasern) verwenden Azetylcholin als Neurotransmitter; die postganglionären Fasern sind entweder cholinerg oder adrenerg.
6. Zusammenfassung
Das PNS umfaßt die 10 Hirnnervenpaare und die 31 Paare von Rückenmarksnerven, sowie alle Ganglien und Rezeptoren, soweit sie außerhalb des ZNS liegen. Es repräsentiert damit den nervösen Leitungsapparat.
Als Rezeptoren bezeichnet man "Empfangseinrichtungen" für bestimmte Reize. Sie werden nach dem Reizursprung im inneren oder äußeren Milieu, nach ihrer Lage im Organismus und nach der Art des zu registrierenden Reizes unterschieden.
Die Wechselwirkung zwischen Reiz und Rezeptorendigung (Transduktion) ist an die Struktur der Zellmembran gebunden und löst eine lokale Potentialverschiebung aus.
Bei den Mechanorezeptoren werden freie von eingekapselten Nervenendigungen unterschieden.
Die wichtigsten Rezeptoren des motorischen Systems sind die Muskelspindeln und Sehnenorgane.
Der allgemeine Aufbau der Rezeptortypen zeigt folgende gemeinsame Merkmale:
- Polare Struktur,
- Keine Myelinhülle am Nervenende,
- Spezialisierungen am Nervenende.
Bei den umkapselten Nervenendigungen treten weitere Merkmale hinzu:
- Hilfsapparate nicht-nervöser Art,
- Einbeziehung von Hilfszellen (z.B. Muskelzellen, sekundäre Sinneszellen, Kapseln),
- Integration in das umgebende Gewebe,
- Lage- und Verteilungscharakteristik.



